
Significance
Many organisms exhibit useful micro- and nanoscale supplies with structural definition and efficiency that problem artificial fabrication methods, but we all know little in regards to the processes that allow their formation. Utilizing butterfly scales as a mannequin system for useful biomaterials, we set up a timeline of scale formation and quantify related structural parameters for growing painted girl butterflies. We overcome challenges of earlier efforts by imaging construction formation instantly in residing organisms, which permits us to constantly observe the evolving wing tissue and the wonderful particulars of particular person scale cells. Visualization of scale construction formation in reside butterflies types the premise for modeling the underlying biomechanical processes and opens avenues for his or her translation into superior fabrication methods.
Summary
Throughout metamorphosis, the wings of a butterfly sprout tons of of 1000’s of scales with intricate microstructures and nano-structures that decide the wings’ optical look, wetting traits, thermodynamic properties, and aerodynamic habits. Though the useful traits of scales are well-known and show fascinating in varied functions, the dynamic processes and temporal coordination required to sculpt the scales’ many structural options stay poorly understood. Present information of scale progress is primarily gained from ex vivo research of mounted scale cells at discrete time factors; to totally perceive scale formation, it’s important to characterize the time-dependent morphological modifications all through their improvement. Right here, we report the continual, in vivo, label-free imaging of rising scale cells of Vanessa cardui utilizing speckle-correlation reflection part microscopy. By capturing time-resolved volumetric tissue information along with nanoscale floor peak data, we set up a morphological timeline of wing scale formation and acquire quantitative insights into the underlying processes concerned in scale cell patterning and progress. We determine early variations within the patterning of canopy and floor scales on the younger wing and quantify geometrical parameters of rising scale options, which counsel that floor progress is important to construction formation. Our quantitative, time-resolved in vivo imaging of butterfly scale improvement supplies the inspiration for decoding the processes and biomechanical ideas concerned within the formation of useful buildings in organic supplies.
The useful buildings of butterfly wing scales kind throughout pupal improvement: scale cells develop protrusions that function templates for finely sculpted nanoscale cuticle morphologies (1⇓–3). By tailoring these scale morphologies, butterflies produce distinctive visible appearances (4⇓⇓–7), guarantee thermal regulation (8) and water repellency (9), or generate useful acoustic (10) and aerodynamic results (11). Interdisciplinary curiosity in these materials functionalities has led to scientific advances within the complete evaluation of the scales’ multifunctional materials properties (12), design of next-generation bioinspired useful supplies (13, 14), identification of key genes in patterning and structural coloration (15⇓⇓⇓–19), and analysis of the affect of ecological elements on biodiversity (20, 21). Though the enviable performance of butterfly wings relies upon closely on the exact structural structure of the wing scales, little is thought in regards to the dynamics, processes, and phenomena concerned in scale improvement (22).
Every scale on a butterfly’s wing is shaped by a person cell, which secretes a chitinous cuticle that types a single-cell exo- skeleton. In lots of butterflies, these scales are additional organized in rows of alternating cowl and floor scale morphologies (1). The mature scales of the painted girl butterfly (Vanessa cardui) exemplify a skeletal scale blueprint, which is extensively mirrored in each the easy and the subtle wing scale morphologies discovered throughout Lepidoptera (Fig. 1A–D). Usually, the higher floor of a scale consists of ridges working down its size; these ridges are composed of overlapping lamellae and are linked by crossribs (23). Supporting trabeculae bridge the higher options and decrease scale floor, which is basically a skinny lamina on the order of 100 nm in thickness. The wealthy range of scale morphologies in different butterflies and moths could also be considered modulations of the fundamental buildings discovered on this generic scale structure. Consequently, the easy-to-rear V. cardui is a positive mannequin system for gaining insights into the processes and mechanical phenomena underlying organic formation of useful micro- and nanostructures (15, 24, 25).

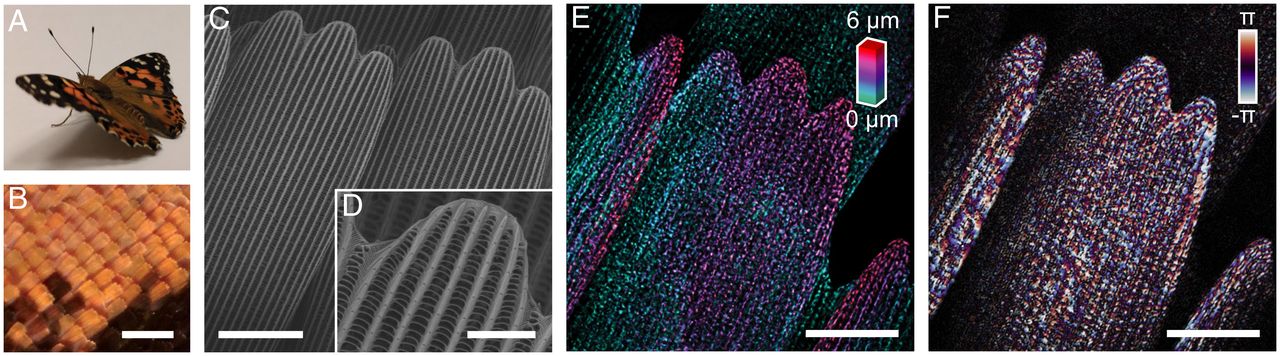
Imaging the construction of totally shaped butterfly scales. (A) The painted girl butterfly, V. cardui. (B) Optical micrograph of orange and black wing scales. (C) Scanning electron micrograph of particular person grownup scales, with ridges working down the size of the size. (D) Magnification of scale finger exhibiting that ridges include stacked lamellae and are linked by crossribs. (E) Volumetric picture of an grownup scale acquired by speckle-correlation reflection part microscopy (crimson, prime of the volumetric information stack; inexperienced, backside). (F) A single slice of part information for a similar scale. (Scale bars: B, 200 µm; C, 20 µm; D, 5 µm; and E and F, 20 µm.)
Key insights into the formation of those buildings have resulted from the evaluation of dissected and stained wing tissues at discrete developmental time factors. Nearly a century in the past, the sequence of mobile division, scale protrusion, progress, and ridge formation was documented in flour moths (26). Since then, electron microscopy served to elucidate nanoscale buildings within the wing tissue and supplied glimpses of cuticle progress on the size cell (27⇓–29). Seminal research emphasised the optical perform of the size and provided hypotheses for lamella formation on the size ridges by way of mechanical wrinkling (30) and for three-dimensional (3D) construction formation by way of inside membrane templates (23). This rationale has been used to clarify how gyroids and different cubic buildings kind in scales (31). Extra not too long ago, confocal imaging has allowed nearer examination of fabric distribution, albeit in mounted wing tissues as a consequence of a scarcity of endogenous labeling strategies in Lepidoptera. These research described the signaling elements concerned in patterning scale positions (32), explored the function of actin within the formation of scale fingers and ridges (24), and quantified actin bundle spacing and chitin distribution throughout and after improvement (25). Along with discrete snapshots from wings mounted at completely different developmental phases (33), the cuticle buildings of mature scales could trace at their formation: in some grownup scales, inside gyroids are ordered in growing measurement up the size of the size, presumably illustrating the timing in onset and progress (34).
Whereas these time-discrete imaging efforts present glimpses into scale improvement, a complete understanding of the processes underlying scale construction formation might be gained solely by steady statement of the spatiotemporal development of residing scale cells (22). Current exogenous fluorescent imaging of reside lepidopteran pupae captured the younger scale cell and the preliminary protrusion of scales (35, 36). Regardless of this progress, visualizing subcellular options of reside scale cells all through improvement stays an unsolved problem, as a consequence of issues inherently related to the imaging of tissues that characteristic heterogenous and pronounced micro- and nanoscale refractive index variations. Moreover, imaging over lengthy durations with fluorescent methods is prone to photobleaching and photodamage; furthermore, genetic constructs for fluorescent labeling in reside organisms are nonetheless restricted for butterflies.
Right here, we report the continual, in vivo, label-free imaging of growing scale cells within the residing V. cardui butterfly utilizing speckle-correlation reflection part microscopy (see Fig. 1 for comparability with scanning electron microscopy information). This quantitative part imaging method gives a flexible technique for observing the expansion of useful supplies in vivo with excessive temporal and spatial decision. We seize important moments of lepidopteran scale construction formation in residing organisms on a steady timeline. Specifically, we recognized a two-step strategy of tissue patterning within the early epithelial sheet and quantified the morphological modifications occurring throughout varied size scales as scale cells develop. Insights from steady imaging of scale formation kind the inspiration for understanding the biomechanical processes concerned within the genesis of useful cuticle morphologies.
Visualization of Scale Options in Dwell Pupae
To picture butterfly wing scales throughout metamorphosis, we developed a specimen preparation and upkeep protocol that allows in vivo quantitative part imaging (Fig. 2). We acquire optical entry to the wing tissue throughout improvement by changing a part of the pupa’s cuticle with a glass window, exploiting quite a lot of surgical methods (Fig. 2 A and B; see Materials and Methods and ref. 35 for timing and methods). Even macroscopically, wing tissue improvement might be monitored via the statement window. Initially, the wing is a translucent epithelial sheet; because the scales produce chitinous cuticle, the wing turns into reflective and ultimately exhibits mature pigment patterns (Fig. 2 B–E). For long-term quantitative observations with excessive spatial decision, the optically uncovered wing is imaged via the window within the chrysalis utilizing speckle-correlation reflection part microscopy (37, 38) (Fig. 2F and SI Appendix, Fig. S1). This interferometric method yields each part and amplitude information from reside and unlabeled specimens, with wonderful axial sectioning, rejecting out-of-focus data (Fig. 2G andI; SI Appendix, Fig. S2; and Movie S1). The amplitude information seize modifications in refractive index, that are normally related to materials interfaces (Fig. 2G). By scanning via the tissue depth, a three-dimensional picture of reside, growing scales and wing tissue might be reconstructed for volumes of 75 × 75 × 200 µm3 with a most lateral decision of 490 nm and a most axial decision of 1.03 µm. We visualize this 3D volumetric amplitude information by color-coding every information slice in line with its peak within the picture quantity (Fig. 2H and Movie S2). The part information seize the peak of the fabric interfaces inside every optical slice (SI Appendix, Fig. S2). The orientation of the part gradient, which encodes the native slope on the scale floor, reveals important scale options (Fig. 2I). Particular person line profiles quantify the size floor peak with sensitivity to variations on the ∼10-nm scale, offering quantitative insights into ridge and lamellae formation all through scale improvement (Fig. 2J and K).

Optical window and in vivo microscopy to picture growing scales. (A) Creating an optical window into the chrysalis: lifting a small flap of cuticle and forewing, revealing the hindwing, and sealing of the uncovered space with a coverslip and a biocompatible adhesive (blue). (B) The uncovered wing at 2% of improvement. The crimson asterisk marks the placement in A. (C–E) Optically accessible wing space at 3%, 73%, and 100% of improvement (minutes earlier than eclosion). (F) Schematic diagram of the in vivo imaging setup. QWP, quarter wave plate; PBS, polarized beam splitter. (G) A single slice of deconvolved mirrored gentle amplitude information, exhibiting the information of forewing scales in a pupa at 83% improvement (8.12 d for this specimen). (H) Volumetric amplitude stack exhibiting overlapping scales (crimson, highest slice; inexperienced, lowest slice via a 6-µm depth). (I) Visualization of micro- and nanoscale options on scales by way of the part gradient related to the amplitude information in G. The coloured cone signifies the lean orientation (crimson, southward; blue, northward downhill slope). (J and Ok) Floor profiles alongside strains indicated in G exhibiting ridges (J) and peak of ridge lamellae (Ok). (Scale bars: B–E, 1 mm; G–I, 20 µm.)
A Phenomenological Timeline of Scale Cell Improvement In Vivo
We observe the event of particular person specimens from the primary few hours after pupation till the organism begins to eclose; the period of the pupal stage (100% improvement) is often about 10 d and may differ by as much as 2 d, with developmental timings depending on the kind of scale and placement on the wing (24, 39). We observe the wing tissue because it transforms from a easy folded epithelial monolayer to a mature wing with totally shaped scale buildings (Fig. 3; Movie S3; and SI Appendix, Fig. S3). At round 1% of improvement, morphologically homogeneous epithelial cells are tightly packed on the wing floor (Fig. 3A); they’re extra sparsely linked on the inside by way of an evolving community of intercellular junctions (SI Appendix, Fig. S4 and Movie S4) which have been beforehand described as ft and cytonemes (36, 40). Mitoses of generalized epidermal cells are readily seen occurring in-plane with the tissue (Movie S4). Then, choose cells–the size precursor cells–swell in measurement (crimson areas in Fig. 3B). These cells bear two divisions: after the primary division, one daughter cell degenerates, whereas the opposite continues to the second division to supply a scale cell and a socket cell (SI Appendix, Fig. S5) (26). The dimensions cell protrudes via the socket, away from the wing floor (Fig. 3C). Though the membrane of the protrusion is initially tough–doubtless as a consequence of microvilli that underlie epicuticle formation (27, 29)–the floor quickly turns into clean (SI Appendix, Fig. S6). The dimensions then expands (Fig. 3D), finally reaching its remaining size and width earlier than 60% of pupal improvement. As the size reaches its remaining size, the forefront splits into fingers and the longitudinal construction of ridges seems (Fig. 3E). In earlier research, scanning electron microscopy has proven that procuticle–a matrix of chitin and protein–grows at these ridge places (27, 29). The ridges turn out to be extra outlined and develop lamellae, whereas the decrease lamina step by step expands to cowl all however the edges of the decrease floor (Fig. 3F).

Steady imaging of scale progress on a person V. cardui pupa. (A) Wing tissue at 5% of pupal improvement, exhibiting cell division previous to precursor cell differentiation. (B) Enlarged, raised precursor cells (crimson, encircled by dashed line) are identifiable at 15% improvement. (C) Scales (two are emphasised by the dashed outlines) have begun to develop away from the sockets on the wing (one is emphasised by the dotted define) at 34% improvement; cowl scales (arrows) and floor scales (arrowheads) are simply distinguishable. (D) Increasing scales and the start of finger formation at 44% improvement. (E) Scales at their remaining measurement with well-pronounced fingers at 62% improvement. Longitudinal buildings are faintly seen. (F) Scales present a decrease lamina (inexperienced zone) beneath developed ridges (crimson zone) at 99% improvement. (G) Timeline of great occasions noticed in growing tissue, permitting for variation between wing places. Proven beneath G is the consultant coloration bar indicating volumetric picture depth for all photos: 0 µm to six µm (A, B, and D–F) and 0 µm to 16.8 µm (C). (Scale bars: A and B, 10 µm; C–F, 20 µm.)
Scale-Forming Processes Implicated by Morphological Dynamics
By monitoring gradual modifications in scale progress over time, we are able to explicitly outline the timing of particular developmental occasions inside particular person pupae and start to grasp the processes guiding morphogenesis (Fig. 3G). Right here, we handle two facets of wing scale improvement: spatial patterning of the size precursor cells within the wing tissue and evolution of scale ridge spacing and peak throughout scale progress.
Scale Precursor Cells of Cowl and Floor Scales Differentiate in a Row at Two Separate Occasions
In V. cardui and different butterflies, scales are organized in neat rows, with alternating floor and canopy scales. Step one on this patterning has been attributed to lateral inhibition by way of the Notch signaling pathway, which results in unfastened rows of remoted precursor cells. This attribution is predicated on a comparability with Drosophila bristle improvement, the place Notch signaling between neighboring cells creates a suggestions loop that leads to spatially remoted, low-Notch cells that turn out to be bristle precursor cells (32, 41, 42). Likewise, within the wings of younger lepidopteran pupae, cells with low Notch expression have been noticed in a unfastened row-like sample (32). The way by which scale precursor cells are organized into their remaining neat row sample has not but been decided, though cell migration and rearrangement have been proposed as hypotheses (32, 40). Little or no is thought about when and the way a precursor cell transitions to a floor or a canopy scale.
Our information in V. cardui present {that a} subset of epithelial cells differentiates into unfastened rows of enormous, remoted precursor cells (Fig. 4A), in settlement with earlier observations of early morphological differentiation (32, 39, 40). Though these precursor cells are initially solely roughly aligned, they subsequently shift into extra outlined rows (Fig. 4B and C) with out touching one another. Then, the smaller cells which might be positioned between the precursor cells additionally differentiate, rising to the big precursor cell measurement (Fig. 4C and D and Movie S5). Because the first and second units of precursor cells alternate down a given row, every group should give rise to both cowl or floor scales. As soon as the scales develop and the shapes of floor and canopy scales are distinguishable, we are able to observe them again in time to find out their relative positions. The distal place and late look of the second set counsel that they’re floor scale precursor cells, and the primary set are the quilt scale precursors (SI Appendix, Fig. S7).

Scale precursor cells seem in two phases. Cowl scale precursor cells (raised crimson spheroids) are labeled 1, 2, and three and are tracked over 13 to 18% of improvement in a single pupa. (A) Precursor cells initially align roughly in rows. (B) Identical precursor cells barely shifted. (C) A brand new precursor cell (arrowhead “a”) begins to enlarge between cell 2 and cell 3; all of the precursor cells transfer towards larger alignment. (D) New floor scale precursor cells (arrowheads) are actually of comparable measurement to the older ones; all precursor cells are organized in a straight row. (A’–D’) Schematic depiction of the precursor cells (magenta). Notice that borders of the encircling cells (inexperienced) usually are not clearly discernible all over the place within the information however are drawn just for illustrative functions. Volumetric picture depth: 0 to six µm. (Scale bar: 20 µm.)
Thus, the formation of tight rows of scales is primarily enabled by the differentiation of a second set of scale precursor cells that spatially alternate in keeping with the primary set. What determines the spatial positioning and timing of differentiation for this second set of precursor cells? Within the improvement of Drosophila bristles—which have regularly been described as homologous to lepidopteran scales (24, 32)—lateral inhibition habits of Notch dynamics can generate short-term stripe patterns of contiguous cells evolving right into a row of nonadjacent bristle precursor cells (42). But our information present that the primary group of scale precursor cells is already nonadjacent in V. cardui, just like the Notch sample on Heliconius erato butterflies (32).
Scale Ridges Develop at Fixed Spacing
To determine geometric constraints and infer biomechanical processes that affect the morphological improvement of scales, we tracked and quantified the evolution of assorted structural parameters in vivo in particular person butterflies (Fig. 5; notice that the exact time of scale improvement could differ, and the scales in Fig. 5 developed barely before these proven in Fig. 3 however adopted the identical development of occasions). A comparatively brief window of time, roughly 35 to 40% of improvement (round 100 h to 114 h), comprises many key moments of scale formation: the size reaches its maximal measurement, fingers kind, and ridges seem (Fig. 5A and B). Scale size and width develop on completely different timelines, with the size retracting barely even earlier than the size reaches its terminal width (Fig. 5C and D). Throughout a lot of scale growth, the already-slender scale step by step continues to skinny (Fig. 5E). But, regardless of this anisotropic progress, estimates of scale quantity and floor space develop in lockstep (Fig. 5F).

Quantification of scale form evolution and ridge progress. (A) Scale morphology at 35% of pupal improvement. Volumetric information (Prime) present midsized scales. Part information (Center) from the area marked within the volumetric information and a peak profile (h) of the part information (Backside) taken alongside the white dashed line point out a comparatively clean scale floor. (B) Scale morphology at 41% of pupal improvement. Scale fingers are obvious in volumetric information. Part information (Center) from marked area point out common spacing of proto-ridges, that are about 100 nm in peak (backside profile extracted from dashed line). (C–F) Measured and imply scale form parameters throughout improvement: size (C), width (D), thickness (E), and quantity (crimson) and floor space (black) as share of most estimated from the bounding field of scales (F). Shaded time spans are laid out in H. (G and H) Ridge parameters throughout improvement: spacing between forming ridges for a number of scales (G). Every curve of linked information factors exhibits steady measurements of a person scale. (H) Characterization of ridge peak (purple) profiles and ridge periodicity (teal). Bars point out SD; curve outlines qualitatively illustrate normal characteristic improvement. White background marks the time span when the size floor is moderately clean; darkish grey shading signifies the time span the place forming buildings are irregular and spacing distribution varies; gentle grey signifies the time span when periodicity of the ridges is decided and peak grows. Information in A–F and H have been acquired from a single pupa; for G a distinct pupa was used to observe ridge improvement on single scales over longer durations of time. Error bars correspond to measurements from three scales. Volumetric picture depth: 0 to six µm. (Scale bars: A and B, Prime, 20 µm; A and B, Center, 10 µm.)
At ∼39% of improvement, lengthy spars of fabric working down the size of the size turn out to be seen within the amplitude information (Fig. 5G). Because the part information present a clean scale floor when these options first seem, these spars are doubtless bundles of actin that can ultimately template the ridges, as has been described beforehand (24, 25, 28). As the size widens, the spacing of actin bundles additionally widens till it reaches ∼1.8-µm separation. Quickly after, the floor of the membrane is not clean, and it turns into troublesome to find out whether or not the longitudinal striations within the amplitude information are the actin serving as ridge templates or the ridges themselves. Earlier work means that actin bundle spacing and cuticular ridge spacing are tightly associated and seem to not change from 40 to 95% improvement (25). We tracked this spacing on particular person scales in our steady information and confirmed that after the size floor is not clean, the periodic spacing certainly stays fixed on particular person scales via the overwhelming majority of ridge improvement (for the measured scales, a minimum of till 89% of pupal improvement).
Our part information reveal how floor morphology modifications on the onset of ridge formation (Fig. 5H and SI Appendix, Fig. S8). The epicuticle on the size floor is clean throughout a lot of the scale growth. The onset of ridge formation seems to start out close to the tip of the scales’ growth (∼37% of improvement for this specimen), after the size has reached its maximal size however earlier than it has attained its full width. Undulations in floor peak seem, however with irregular spacing and ranging distribution throughout the size’s floor. This spatial and temporal heterogeneity could also be as a consequence of variations within the onset of cuticle manufacturing. Then, at ∼39% improvement, periodicity turns into extremely common and stays common because the nascent ridges develop in peak. Part and amplitude information collectively point out that ridge spacing stays almost fixed from the earliest moments of ridge look and progress till late in scale improvement, when a lot of the ridges’ wonderful construction has been established; we’ve not but decided whether or not there are any spacing modifications after 89% improvement via eclosion and the drying of the scales.
Different research have proven that ridges seem between repeatedly spaced bundles of actin filaments (25, 28), that are mandatory for correct ridge formation (24). As ridges develop, they develop their wonderful lamellar construction. One speculation, established nearly 40 y in the past, proposes that lamellar formation is a wrinkling phenomenon pushed by mechanical buckling of the cuticle layer with the required stress arising from a lower in distance between the actin bundles (30).
Nonetheless, our information point out that ridge spacing stays fixed after preliminary ridge look, implying {that a} lower in actin bundle spacing is unlikely and thus can’t drive ridge formation. However, buckling stays a believable mechanism for the formation of ridges and lamella, doubtlessly ensuing from progress of the size floor topic to an underlying mechanical constraint. The rising floor space of membrane and deposited cuticle could also be spatially constrained by the actin bundles, resulting in out-of-plane buckling; alternatively, differential progress between the membrane and cuticle may result in stresses that induce ridge look (43). Additional research are wanted to disclose the biomechanical phenomena underlying the interaction of actin, cell membrane, and deposited cuticle that drive the formation of the size ridge buildings.
Conclusion
Utilizing speckle-correlation reflection part microscopy and lepidopteran surgical methods, we exhibit label-free steady visualization and quantitative characterization of microscale construction formation in reside lepidopterans, gaining quantitative insights about mobile group and improvement of subcellular options on single scales all through pupal improvement. On the tissue degree early in improvement, we noticed the sequential patterning of precursor cells for canopy and floor scales. Our observations increase questions in regards to the signaling that allows floor scale differentiation, since earlier work signifies that the primary set of precursor cells displays low Notch and represses scale-forming transcription elements in its neighbors (32, 42), which incorporates the second set of precursor cells. Additional research are required to grasp how the bottom scale sample is organized between the quilt scales. On the single-cell degree, our quantification of the spacing and peak of scale ridges reveals that the ridge spacing stays fixed from the primary incidence of ridges at about 47% improvement to a minimum of 89% improvement. This means that the formation of ridges is unlikely to end result from a discount in actin bundle spacing as hypothesized in earlier work (30), but could also be pushed by floor progress and buckling when constrained by the actin bundles. Future research with our imaging method could additional characterize scale materials progress and inform quantitative mechanical fashions which might be wanted to establish the biophysical mechanisms underlying the formation of the scales’ wonderful ridges and crossribs. We anticipate that our imaging technique could speed up developmental research in different species of Lepidoptera and even different orders, because it bypasses the necessity to develop endogenous labeling or sustained exogenous labeling. The development of scale construction formation in V. cardui is prone to have important parallels to different lepidopterans. A comparability between the size buildings of various species and their respective improvement timelines will assist determine the processes answerable for creating and fine-tuning structural options. Future work on pushing the boundaries of decision of the offered part imaging method, quantitative mapping of refractive index distribution, and standardization of key system parts could facilitate the implementation of our method in different research of organic materials formation and mobile and tissue improvement. Moreover, our in vivo imaging could also be used at the side of genome enhancing protocols (15, 16) or molecular inhibition (24) to elucidate genetic and molecular mediators of construction formation. We take into account label-free quantitative part imaging on entire residing organisms a vital software within the quest for shedding gentle on the interaction of genetics, proteomics, and biomechanics to allow construction phenotypes with particular performance. Insights into the formation ideas of the multifunctional materials buildings employed by bugs may permit us to glean particular manufacturing methods with management over materials structure throughout all functionally related size scales.
Supplies and Strategies
Butterfly Rearing
Painted girl (V. cardui) butterflies have been raised in our laboratory with unique inventory obtained from Carolina Organic Provide Firm. Larvae are monitored in particular person containers to determine the time of molting of the larval pores and skin to inside 15 min. The share of improvement of a person after surgical procedure is estimated utilizing the common period of pupal improvement of different pupae from the identical era. Further particulars are included in SI Appendix.
Pupal Surgical procedure
We use two surgical approaches to reveal the forewing and hindwing tissue within the chrysalis, respectively. To show the forewing for single time level imaging (Fig. 2A), we take away a part of the cuticle from above the forewing. This cuticle might be eliminated at any time after apolysis, which happens at ∼20% of improvement (SI Appendix, Fig. S3). We anesthetize the pupa in a small chamber that has been flushed with carbon dioxide for five to 10 min to immobilize the organism whereas making a small (∼1 × 1 mm), shallow reduce within the chrysalis with a scalpel (Feather; Micro Scalpel) to take away the cuticle. We then place a 150 µm-thick (VWR; no. 1) glass coverslip over the uncovered tissue and seal it with a light-curing dental composite (Pentron; Circulation-It ALC) utilizing a handheld dental curing lamp (NSKI; LY-02 LED Mild Remedy Lamp).
To show the hindwing for steady imaging, we tailored the surgical technique described by Otaki and coworkers (35, 44). We grasp the chrysalis cuticle of a not too long ago molted pupa along with the forewing and fold it anteriorly towards the pinnacle (these youthful pupae don’t require immobilization). To restrict the unfold of any potential melanization immune response, we separate the hindwing from the forewing with a strip of dental composite. Once more, we positioned a glass coverslip over the uncovered wing and sealed it with dental composite. Further particulars are included in SI Appendix.
Speckle-Correlation Reflection Part Microscopy
We make use of the interferometry method beforehand revealed (37, 38) (SI Appendix, Fig. S1). Briefly, a supercontinuum laser (NKT Photonics; SuperK Excessive EXR-4) is shipped via a rotating diffuser to supply a dynamically various speckle illumination. With a bandpass filter, heart wavelength 800 nm is chosen with 40 nm bandwidth. The illumination is then distributed with a polarizing beam splitter to the goals (Olympus; LUMPLFLN 60XW, 1.0NA water dipping) on the reference arm and the pattern arm; quarter-wave plates trigger the reference and pattern sign polarizations to stay orthogonal to one another. Off-axis holography is achieved by way of a grating to divide the mixed alerts, adopted by a mixture of polarizers and quarter-wave plates to filter and align the alerts for interference. The interferogram is then captured by a digital camera (Level Grey; Flea3) with as much as 100 fps pace and processed in Fourier area to collect the amplitude and part data from the pattern. A motorized stage within the pattern stage scans the vertical place of the pattern to accumulate a volumetric picture. The theoretical lateral decision of 490 nm and the theoretical axial decision of 1.03 µm have been beforehand corroborated with measurements (37). When imaging beneath a floor in vivo, the decision is anticipated to be decrease as a consequence of a number of scattering and spherical aberration from the tissue and doubtlessly may step by step degrade over time if the reference arm drifts.
Imaging Protocol
We sometimes start to picture the pupa inside 1 h to three h after surgical procedure. We place the pupa beneath the reference arm of the speckle-correlation reflection part microscope and scan alongside the optical axis (z axis) in 400-nm steps to construct up a quantity of picture slices. Acceptable imaging depth is algorithmically decided by finding the depth of most area-integrated depth. The reference arm is adjusted to match the change in optical path size. A coverslip with a reflective gold coating on the far aspect is used because the reference mirror to cancel the dispersion and path-length delay launched by the pupa’s coverslip.
For steady imaging, a program mechanically cycles via the focusing and picture acquisition course of each 15 min, with microscope alignment checked in individual as soon as per day; a syringe pump periodically replaces water misplaced by evaporation on the immersion lenses. For single time level photos (e.g., Fig. 2G–J), carbon dioxide is flowed over the pupa for five min to make sure immobilization instantly previous to imaging. Computation and interface management have been executed utilizing MATLAB (MathWorks). Further particulars are included in SI Appendix.
Information Processing and Visualization
We discovered that 10 iterations of Lucy–Richardson deconvolution with the theoretically estimated level unfold perform (45) are efficient for visualizing scale options within the amplitude information, notably in middle- to late-stage scale buildings (Figs. 1 and 2).
The colour representations of 3D amplitude information are based mostly on the Temporal-Shade Code plugin by Kota Miura for Fiji/ImageJ. Picture slices are coloured in line with their depth place, after which a mean depth is taken alongside the quantity depth for every RGB channel.
The preliminary part information are wrapped on , which might be described as
[1]the place W is an operator for wrapping, given by
[2]
with as an integer that ensures wrapping; i.e., . Itoh (46) described the unwrapping course of for factors of wrapped part information in a single dimension:
[3]
the place Δ is an operator for the differential, . We use this method to investigate 1D profiles in our part information.
In 2D, unwrapping can endure from the results of noise and cliffs, that are compounded when contemplating each dimensions (47). We characterize the native part habits by way of the integrand of the unwrapping scheme,
[4]which represents the native part gradient alongside a selected dimension. Mixture of the horizontal and vertical parts of this gradient permits us to find out the route of slope of the floor. This visualization technique permits discernment of wonderful options within the midst of a lot bigger depth modifications. Further particulars are included in SI Appendix.
Picture Evaluation
We use Dragonfly (Object Analysis Techniques) to measure size, width, and thickness of canopy scales in deconvolved 3D amplitude information (SI Appendix, Fig. S9). Size was extrapolated for scales that lay partly outdoors the imaging window (SI Appendix). Floor space and quantity are estimated utilizing the measured size, width, and thickness to outline the presumptive bounding field of the size.
To measure ridge peak and spacing within the part information (SI Appendix, Fig. S10), we unwrap and rotate a 1D profile from a slice of part information. We discover the dominant spatial frequency by way of Fourier evaluation. We outline the peak of the height as the space from the height to the road connecting the troughs on both aspect of the height. The imply and SD peak peak are calculated for every profile; all profiles are then used to find out the pooled imply and the pooled SD for every time level.
To measure the actin/ridge spacing within the amplitude information (SI Appendix, Fig. S11), we first rotate the info to seize the aircraft of a scale and outline the area of curiosity of a scale in Dragonfly. We then discover the dominant spatial frequency by way of 2D Fourier evaluation. Further particulars are included in SI Appendix.
Acknowledgments
We thank Youngwoon Choi (Korea College) for insights on speckle-correlation reflection part microscopy, Yong Zhang (Massachusetts Institute of Know-how [MIT]) for assist with gold-coating coverslips, Bodo Wilts (College of Fribourg) for feedback on the manuscript, Cindy Lu for discussions on cell differentiation, and Meera Singh (MIT) and Julia Kudryashev (MIT) for help in elevating butterflies and early surgical trials. This work was supported by the Nationwide Science Basis via the “Designing Supplies to Revolutionize and Engineer our Future” program (DMREF-1922321) and CBET program on “Particulate and Multiphase Processes” (Grant 1804241) (to A.D.M. and M.Ok.). S.Ok., Z.Y., and P.T.C.S. acknowledge assist by NIH Grants P41EB015871, R21GM140613, R01HL158102, and R01DA045549. P.T.C.S. additional acknowledges assist by Grant U01CA202177, Division of Vitality DE-FOA-0002359; Hamamatsu Company; Samsung Superior Institute of Know-how; Singapore–MIT Alliance for Analysis and Know-how Middle–Crucial Analytics for Manufacturing Customized-Medication Interdisciplinary Analysis Group (SMART CAMP IRG); HARDi Holdings Restricted; Fujikura Ltd.; and Hong Kong Innovation and Know-how Consortium.
Footnotes
- Accepted October 11, 2021.
-
Creator contributions: A.D.M., S.Ok., Z.Y., P.T.C.S., and M.Ok. designed analysis; A.D.M. and S.Ok. carried out analysis; A.D.M., S.Ok., Z.Y., P.T.C.S., and M.Ok. contributed new reagents/analytic instruments; A.D.M., S.Ok., and M.Ok. analyzed information; A.D.M. and M.Ok. wrote the paper; and A.D.M., S.Ok., Z.Y., P.T.C.S., and M.Ok. mentioned and reviewed the manuscript.
-
The authors declare no competing curiosity.
-
This text is a PNAS Direct Submission.
-
See online for associated content material reminiscent of Commentaries.
-
This text comprises supporting data on-line at https://www.pnas.org/lookup/suppl/doi:10.1073/pnas.2112009118/-/DCSupplemental.
- Copyright © 2021 the Creator(s). Revealed by PNAS.













{kind=link}