
Significance
Migratory locusts show putting phenotypical plasticity. Gregarious locusts at excessive density can migrate lengthy distances and trigger big financial losses of crops. In contrast, solitary locusts at low density have restricted skill in long-distance flight. Nevertheless, the mechanisms underlying such flight capability variation are poorly understood. Right here, we discovered that the flight muscle of solitary locusts has a better catabolic capability that’s related to better reactive oxygen species (ROS) technology throughout high-velocity flights. In contrast, a comparatively decrease catabolic capability in gregarious locusts is related to decrease ROS technology throughout long-distance flights. This discovering uncovers the metabolic mechanism of locust flight trait alteration in response to density modifications and enhances our understanding of the organic processes enabling locust migration.
Summary
Flight skill is crucial for the big variety and evolutionary success of bugs. The migratory locusts exhibit flight capability plasticity in gregarious and solitary people carefully linked with completely different density experiences. Nevertheless, the differential mechanisms underlying flight traits of locusts are largely unexplored. Right here, we investigated the variation of flight capability by utilizing behavioral, physiological, and multiomics approaches. Behavioral assays confirmed that solitary locusts possess excessive preliminary flight speeds and short-term flight, whereas gregarious locusts can fly for an extended distance at a comparatively decrease velocity. Metabolome–transcriptome evaluation revealed that solitary locusts have extra lively flight muscle power metabolism than gregarious locusts, whereas gregarious locusts present much less proof of reactive oxygen species manufacturing throughout flight. The repression of metabolic exercise by RNA interference markedly decreased the preliminary flight velocity of solitary locusts. Elevating the oxidative stress by paraquat injection remarkably inhibited the long-distance flight of gregarious locusts. In respective crowding and isolation therapies, power metabolic profiles and flight traits of solitary and gregarious locusts have been reversed, indicating that the differentiation of flight capability trusted density and will be reshaped quickly. The density-dependent flight traits of locusts have been attributed to the plasticity of power metabolism and diploma of oxidative stress manufacturing however not power storage. The findings supplied insights into the mechanism underlying the trade-off between velocity and sustainability in animal locomotion and motion.
Insect flight capability is dependent upon a number of elements, akin to wing morphology, hormone regulation, and power metabolism (1). Plastic modifications in wing morphology have been intensively studied in wing-dimorphic bugs (2⇓–4). Juvenile hormone, ecdysteroids, and insulin or insulin-like development issue signaling pathways contribute to wing dimorphism regulation (5⇓–7). In contrast to wing-dimorphic bugs, wing-monomorphic bugs can remodel their flight capability inside one technology (8). A central problem in learning wing-monomorphic insect flight is knowing how bugs regulate power metabolism for a long-distance flight given their small physique dimension and comparatively brief life span (9). Animal flight is among the many bodily actions with the very best power consumption. Throughout flight, the metabolic price (MR) is no less than 50 occasions that in regular bodily actions (10, 11). A hyperlink between gasoline power metabolism and dispersal skill has been instructed in some butterfly species, i.e., the diploma of dispersal will increase with the MR (12, 13). Nevertheless, the connection between flight MR and flight capability stays controversial.
Locusts are probably the most harmful agricultural pests on this planet. The migratory locust (Locusta migratoria) is a typical wing monomorphic insect that displays reversible part modifications between solitary and gregarious kinds in response to inhabitants density variation (14, 15). The morphology, physiology, and behavioral traits of gregarious locusts at excessive inhabitants density remarkably differ from these of conspecific solitary locusts at a low inhabitants density (16⇓–18). With rising inhabitants density, gregarious locusts can type extraordinarily dense hopper bands or migratory grownup swarms, inflicting appreciable harm inside a short while. Furthermore, gregarious adults can fly constantly for ∼12 h, inflicting plague outbreaks in distant areas. In contrast, solitary adults possess restricted efficiency in long-distance flight, and so they fly solely when in search of mates and meals or escaping from predators (14). Variations in habits, physique shade, immunity, metabolism, and replica between gregarious and solitary locusts have been more and more elucidated over time (16⇓–18). As a wonderful mannequin system for insect-flight research, the migratory locust has been extensively used to research the energetic mobilization and metabolic regulation of flight (19). Gregarious locusts have greater lipid reserves and better hyperlipidemic responses to flight than solitary ones. The content material of adipokinetic hormones (AKHs), a neuropeptide household that regulates energetic mobilization throughout extended flight, is greater within the corpora cardiac of solitary locusts than that of gregarious locusts (20, 21). Nevertheless, regulatory elements and mechanisms underlying the differentiation of flight capability of gregarious and solitary locusts require additional elucidation.
The event of multiomics know-how gives highly effective instruments for learning insect habits. As an illustration, comparative transcriptomic and genomic analyses have uncovered variations in gene expression between migratory and nonmigratory monarch butterflies or between the jap and western populations of the monarch butterfly (22, 23). A mixture of tethered flight assessments and transcriptomic analyses has been used to find out transcriptional profile variations related to flight exercise in cotton bollworm (24). A genome-wide evaluation of the migratory locust revealed the growth of gene households concerned in power consumption and cleansing, thereby implying the genetic bases of long-distance flight capacities (25). Metabolome evaluation can present a “nearer” glimpse into the phenotypic options of an organism and has been broadly used to disclose the important thing elements of insect organic processes (26). Due to this fact, a mixture of metabolome and transcriptome evaluation is a promising technique to elucidate the metabolic mechanism of the flight capability variation of solitary and gregarious locusts.
Within the current examine, we in contrast the flight traits and metabolomic and transcriptional profiles of solitary and gregarious locusts. Solitary locusts have excessive flight speeds with brief flight durations. In distinction, gregarious locusts are able to long-distance flight at comparatively low velocity. Metabolomic and transcriptomic analyses mixed with purposeful research revealed that the divergence of flight traits in solitary and gregarious locusts was resulting from their variations in power metabolism and diploma of oxidative stress manufacturing. The crowding and isolation of solitary and gregarious locusts reversed the metabolic profiles and behavioral traits, respectively. Thus, the flight trait differentiation of solitary and gregarious locusts was attributed to completely different inhabitants density experiences linked with different power metabolism and oxidative stress manufacturing.
Outcomes
Solitary Locusts Possess Greater Flight Pace and Intermittent Flying, whereas Gregarious Locusts Exhibit Longer Flight Period.
To analyze the flight efficiency we chosen the 14-d-old grownup locusts for flight behavioral assay on computerized flight mills (27). Based mostly on the mathematical mannequin, 4 parameters of locusts, particularly, most and common flight velocity, flight distance, and flight period, have been recorded to replicate the flight efficiency. No important variations in these parameters between men and women have been noticed after 1 h of flight take a look at on the mills (SI Appendix, Fig. S1) no matter locust part standing. To exclude the attainable interference of replica on flight efficiency solely male adults have been used for additional research.
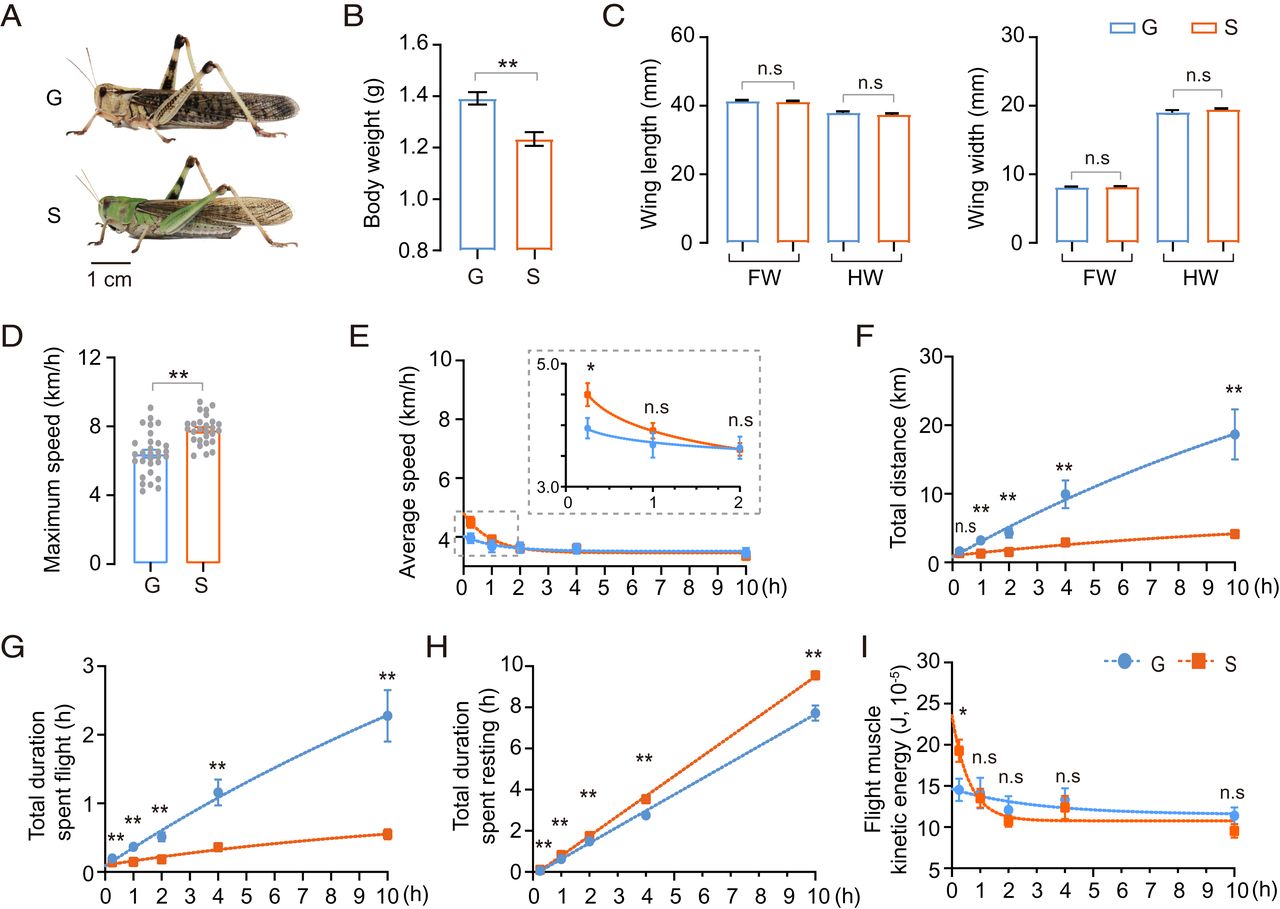
Morphological variations exist between gregarious and solitary locusts (Fig. 1A). Thus, we evaluated the impact of morphological traits on flight efficiency. Gregarious locusts had bigger physique weights than solitary ones (Fig. 1B), however no variations in wing size and wing width have been noticed (Fig. 1C). Correlation analyses confirmed that physique weight had no important correlation with most flight velocity, common flight velocity, flight period, and flight distance, as proven in SI Appendix, Fig. S2.

Flight trait divergence between gregarious and solitary locusts. (A) Typical gregarious and solitary grownup locusts. G and S characterize gregarious and solitary locusts, respectively (n = 28 people, Pupil’s t take a look at). (B and C) Morphological comparisons between G and S. FW and HW characterize forewing and hindwing, respectively (n ≥ 20 people, Pupil’s t take a look at). (D) Comparability of most flight velocity between G and S. Every dot represents a single particular person (Mann–Whitney U take a look at). (E–H) The measurement of flight parameters (E) common flight velocity, (F) complete flight distance, (G) complete period of flight, and (H) complete period of resting. (I) Comparability of flight muscle kinetic power between G and S throughout flight. Flight efficiency for 0.25-, 1-, 2-, 4-, and 10-h assessments have been in contrast. Information for every time level was obtained independently, n ≥ 20 people at every time level (Mann–Whitney U take a look at). The becoming curve was obtained by nonlinear regression. All information are offered because the imply ± SEM, *P < 0.05, **P < 0.01, and n.s, not important.
To analyze the variations in flight efficiency between gregarious and solitary locusts we recorded the flight parameters at 0.25, 1, 2, 4, and 10 h of flight take a look at. On the preliminary stage (0 to 0.25 h), solitary locusts displayed about 25% and 10% greater most (Fig. 1D) and common flight speeds (Fig. 1E), respectively, than gregarious locusts. Nevertheless, the flight velocity of solitary locusts dropped quickly inside 1 h and had no important distinction from that of gregarious locusts in keeping with the outcomes of extended flight assessments (≥1 h) (Fig. 1E). Moreover, throughout extended flight assessments (over 1 h), gregarious locusts exhibited higher efficiency when it comes to complete flight distance and period (Fig. 1 F and G), however solitary locusts confirmed a for much longer complete resting period (Fig. 1 H). Based mostly on common flight velocity and flight muscle mass, flight kinetic power was generated (28). Solitary locusts confirmed considerably greater flight muscle kinetic power at time factors inside 0.25 h. Throughout longer flight assessments (over 0.25 h) there was no important distinction between gregarious and solitary locusts (Fig. 1I). In 10-h assessments on flight mills, the common flight period of gregarious locusts was ∼2.28 h. Some gregarious people have been capable of fly continually for ∼8 h, with a flight distance of over 40 km. In contrast, the common flight period of solitary locusts within the 10-h take a look at was solely ∼0.55 h, and the very best flight distance was not more than 5 km. Thus, solitary locusts flew at excessive velocity just for a short while. Conversely, gregarious locusts have been capable of fly continually at comparatively low velocity for a very long time.
Excessive Degree of Power Metabolism Confers Excessive-Pace and Quick-Time period Flight to Solitary Locusts.
To discover the molecular foundation of flight trait differentiation we performed the transcriptomic evaluation of flight muscle groups in gregarious and solitary locusts. First, gene-expression profiles have been in contrast when the bugs have been at relaxation situations. A complete of 741 differentially expressed genes (DEGs) have been recognized with P < 0.01. Roughly 451 of 741 DEGs have been extremely expressed within the flight muscle groups of solitary locusts and have been enriched largely in pathways concerned in power metabolism, together with oxidative phosphorylation, carbon metabolism, tricarboxylic acid (TCA) cycle, glycolysis, fatty acid degradation, pyruvate metabolism, and fatty acid metabolism in keeping with Kyoto Encyclopedia of Genes and Genomes (KEGG) signaling pathway evaluation (adjusted P < 0.01, Fisher’s precise take a look at). Nevertheless, the 290 DEGs extremely expressed in gregarious locusts confirmed no important enrichment in particular pathways (Fig. 2A and SI Appendix, Fig. S3). Due to this fact, the obvious distinction between gregarious and solitary locusts in flight muscle groups was power metabolism.

Power metabolic profile variations between gregarious and solitary locusts. (A) KEGG enrichment of DEGs. Solely KEGG phrases with adjusted P < 0.01 are proven (Fisher’s precise take a look at). (B) Warmth map representing the gene-expression ranges is concerned in power metabolism underneath resting situations. The 4 numbers on the highest dendrograms characterize the completely different power metabolism processes, as follows: 1, glycolysis and pyruvate metabolism; 2, fatty acid metabolism; 3, citrate cycle; and 4, oxidative phosphorylation. Warmth-map sign signifies log2 fold-change worth relative to the imply expression degree throughout the group. Crimson sign represents greater expression, whereas blue represents decrease expression. (C) Enzyme exercise of hexokinase (HK), pyruvate kinase (PYK), and citrate synthase (CS) (n ≥ 7, Pupil’s t take a look at). (D) Acetyl-CoA and NADH content material measurement within the flight muscle of locusts (n ≥ 7, Pupil’s t take a look at). (E) MR measurement (n = 11, Pupil’s t take a look at). (F) Warmth map representing the expression ranges of genes concerned in power metabolism at 0.25- and 1-h postflight therapy. (G) Comparability of CS enzyme exercise underneath flight situations (n = 7, Pupil’s t take a look at). (H) CS knockdown abolished CS RNA degree and enzyme exercise (n ≥ 7) and decreased CO2 manufacturing price (n ≥ 11, Pupil’s t take a look at). The degrees have been examined 72 h after dsRNA injection. (I and J) Comparability of speed-related flight parameters between controls and CS-silencing locusts (n ≥ 20 people, Mann–Whitney U take a look at). Every dot represents a single particular person. dsCS represents CS knockdown. dsGFP served because the management. Imply ± SEM. Vital variations are denoted by *P < 0.05, **P < 0.01, and n.s, not important. G and S characterize gregarious and solitary locusts, respectively.
To additional authenticate the gene-expression profiles, DEGs have been confirmed by real-time PCR (qRT-PCR). Per transcriptome information (Fig. 2B), the transcription ranges of genes concerned in carbohydrate metabolism—phosphoglucomutase (PGM), hexokinase (HK), pyruvate kinase (PYK), glycogen phosphorylase (GLY), and fructose-bisphosphate aldolase (ALDO)—lipid metabolism—carnitine O-palmitoyltransferase1 (CPT1), carnitine O-palmitoyltransferase2 (CPT2), fatty acid binding protein (FABP), and really long-chain acyl-CoA dehydrogenase (ACADVL), TCA cycle, and oxidative phosphorylation—citrate synthase (CS), cytochrome c oxidase subunit4 (COX4), cytochrome c oxidase subunit7A (COX7A), adenosine 5′-triphosphate (ATP) synthase (ATPsyn), and NADH dehydrogenase subcomplex subunit (NDUFA)—have been greater in solitary locusts than in gregarious ones (SI Appendix, Fig. S4). The actions of three rate-limiting enzymes HK, PYK, and CS have been measured. In step with the gene-expression profiles, the actions of the enzymes have been considerably greater in solitary locusts than in gregarious locusts (Fig. 2C). The degrees of metabolites, acetyl-CoA and NADH, which have been concerned in mitochondrial cardio metabolism, have been greater within the flight muscle groups of solitary locusts (Fig. 2D). Equally, the resting CO2 manufacturing price was considerably greater in solitary locusts (Fig. 2E). Thus, when at relaxation, solitary locusts are extra lively in power metabolism than gregarious ones.
Subsequent, the transcriptome and gene-expression profiles underneath flight situations have been examined. Transcriptomic evaluation and qRT-PCR verification indicated that the power metabolism gene expressions have been considerably greater in solitary locusts than in gregarious locusts in any respect flight-test time factors (flight for 0.25 and 1 h; Fig. 2F and SI Appendix, Fig. S4). Furthermore, the enzyme actions of HK, PYK, and CS have been additionally considerably greater in solitary locusts throughout flight (Fig. 2 G and SI Appendix, Fig. S5). Due to this fact, solitary locusts maintained a better power metabolism throughout flight.
As one of many rate-limiting enzymes of the TCA cycle, CS was assumed to be an necessary management level for metabolic exercise dedication (29). To confirm the impact of power metabolism on the flight traits of the locusts, the metabolic degree was repressed by flattening CS. The injection of double-stranded RNA (dsRNA) of CS considerably repressed the gene expression and enzyme actions in flight muscle groups (Fig. 2H). In the meantime, the knockdown of CS considerably repressed the resting CO2 manufacturing price (Fig. 2H), and the preliminary flight velocity (Fig. 2 I and J), however it had no apparent impact on extended flight efficiency. Due to this fact, this consequence suggests {that a} excessive Krebs cycle capability, as noticed in solitary locusts, is important for the excessive preliminary flight speeds that the solitary-phase locusts exhibit.
Speedy Reactive Oxygen Species Accumulation Impairs the Extended Flight Means of Solitary Locusts.
To match metabolic modifications with gene expression profiles we examined the metabolite dynamics of solitary and gregarious locusts throughout flight. Ultrahigh-performance liquid chromatography–high-resolution mass spectrometry was used to match the relative ranges of metabolites after 0, 0.25, and 1 h of flight therapy. The principal element evaluation (PCA) revealed that the samples of gregarious and solitary locusts have been considerably separated into two classes and the general metabolite distributions have been rather more intense in response to flight in solitary locusts than in gregarious ones (Fig. 3A). Furthermore, 218 and 356 metabolites confirmed important variations in response to flight therapies in gregarious and solitary locusts, respectively (P < 0.05; SI Appendix, Table S1). For all of the differential metabolites, the highest 5 metabolite courses have been discovered to be triacylglycerol (TAG), glycerophosphocholine (PC), glycerophosphoethanolamine (PE), amino acid, and carbohydrate (SI Appendix, Fig. S6). TAG and carbohydrate decreased in the course of the flight of gregarious and solitary locusts, however TAG dropped extra quickly in solitary locusts than in gregarious ones after 0.25 h of flight (Fig. 3B and SI Appendix, Fig. S7). Plasmalogen, a category of glycerophospholipids containing a vinyl-ether and an ester bond on the sn-1 and sn-2 positions, was maintained at a better degree in gregarious locusts (Fig. 3C).

Metabolome evaluation reveals altered oxidative tolerance of solitary and gregarious locusts. (A) PCA trajectory evaluation of altered metabolites in gregarious locusts (blue level) and solitary locusts (orange level) in response to 0.25 and 1 h of flight therapy. Bar traces point out SEM of PC1 and PC2. 5 organic replicates for every flight therapy time level, and every replicate incorporates six locusts. Content material modifications of (B) TAG and (C) PE and PC throughout flight. Warmth-map sign signifies log2 fold-change worth relative to the median expression degree throughout the gregarious and solitary teams, respectively. Yellow and blue alerts characterize greater and decrease ranges, respectively. The dendrograms generated by the hierarchical clustering of metabolites are additionally supplied. TAG: triacylglycerol, PC: glycerophosphocholine; PE: glycerophosphoethanolamine (n = 5 replicates, six locusts per replicate). (D) Pathway topology evaluation of altered metabolites after flight therapy. Solely pathways with P < 0.05 are proven (Fisher’s precise take a look at). (E) Content material modifications in purine intermediate metabolite ranges and (F) GSH throughout flight (n = 5). (G) H2O2 alterations in response to flight (n = 8). Time factors are proven as 0.25- and 1-h flight therapy, every time level is in comparison with relaxation situation samples, Pupil’s t take a look at. (H) GSH/GSSG and H2O2 alterations in response to paraquat therapy within the flight muscle groups (n ≥ 7, Pupil’s t take a look at). (I) Flight efficiency measurement in response to paraquat therapy (n ≥ 17 people, Mann–Whitney U take a look at). Every dot represents a single particular person. Paraquat (2 µL) was injected at concentrations of 4, 20, and 100 mmol⋅L−1 for 48 h. Locusts injected with saline (2 µL) served because the damaging management. Completely different focus samples are in comparison with the damaging management. The info are proven as imply ± SEM, *P < 0.05, **P < 0.01, and n.s, not important. G and S characterize gregarious and solitary locusts, respectively.
Metabolic pathway enrichment was subsequently performed utilizing the overrepresentation methodology. Fisher’s precise take a look at and out-degree centrality algorithm have been used for pathway topology evaluation. The metabolites associated to butanoate, starch, sucrose, alanine, and glycerophospholipid metabolism exhibited related change developments between gregarious and solitary locusts after the flight therapy (Fig. 3D). Along with the widespread pathways that have been enriched in each solitary and gregarious locusts, pathways concerned in purine metabolism, glutathione metabolism, beta-alanine metabolism, and aminoacyl-transfer RNA biosynthesis have been particularly enriched in solitary locusts (P < 0.05; Fig. 3D). The contents of metabolites concerned in purine metabolism (inosinic acid, hypoxanthine, xanthine, and uric acid), and glutathione metabolism, akin to decreased glutathione (GSH) and oxidized glutathione (GSSG), modified considerably in solitary locusts in response to the flight therapy. The relative ranges of uric acid and GSH in solitary locusts decreased considerably after the 0.25-h flight therapy, and their ranges have been low till 1 h of flight. In contrast, the degrees of uric acid and GSH within the gregarious locusts remained unchanged (Fig. 3 E and F).
Uric acid and GSH are necessary antioxidants that quickly lower in solitary locusts, however gregarious and solitary locusts confirmed no important distinction in antioxidant genes expression (SI Appendix, Fig. S8). Thus, we speculated about the potential for speedy reactive oxygen species (ROS) manufacturing. When testing the impact of flight on H2O2 degree in flight muscle groups, we discovered that H2O2 content material dramatically elevated in solitary locusts in any respect time-course factors, however no important enhance was present in gregarious locusts after the identical flight therapy (Fig. 3G).
To discover the impact of oxidative stress on flight efficiency, we elevated oxidative stress by injecting completely different concentrations of paraquat at a dose vary of 0.008 to 0.2 µmol per locust. The contents of H2O2 and GSH/GSSG, in addition to flight efficiency, have been measured. When the focus of paraquat was above 0.04 µmol, a big elevation in H2O2 degree and discount in GSH/GSSG degree was noticed (Fig. 3H). In the meantime, 0.04 µmol of paraquat was enough to repress flight distance and period (Fig. 3I). Thus, the restricted flight period of solitary locusts was presumably as a result of speedy accumulation of oxidative stress.
Gregarious Locusts Keep Extra Energetic Storage.
To analyze whether or not power storage is a determinant of differential flight traits of gregarious and solitary locusts we measured the contents of lipid and glycogen, each of that are necessary power assets in fats our bodies. Lipid reserves are saved primarily as TAG in lipid droplets (LDs) (30). The dimensions of LDs from 14-d-old adults was measured via Nile pink staining. The relative sizes of LDs have been considerably bigger (Fig. 4A) and TAG content material was greater (Fig. 4B) in gregarious locusts. Nevertheless, gregarious and solitary locusts didn’t considerably differ in glycogen content material (Fig. 4B). These information confirmed that gregarious locusts had bigger TAG storage than solitary ones. When solitary and gregarious locusts have been compelled to fly for 1 h, which was roughly twofold the common complete flight period of solitary locusts in 10 h of flight assessments (Fig. 1F), no apparent discount in TAG content material was noticed (Fig. 4C). Thus, the flight period and distance of solitary locusts weren’t restricted by the low power storage in fats our bodies.

Ranges of TAG and glycogen storage. (A) Nile pink staining was carried out in fats our bodies and the relative dimension of LDs was quantitatively analyzed. (B) Relative TAG abundance and glycogen content material measurement (n = 5, Pupil’s t take a look at). (C) TAG content material measurement earlier than and after 1 h of forced-flight therapy (n ≥ 5, Pupil’s t take a look at). G and S characterize gregarious and solitary locusts, respectively. The info are proven as imply ± SEM, **P < 0.01, and n.s, not important.
Inhabitants Density Experiences Set up Primary Power Metabolism and ROS Era for Flight Traits.
To find out whether or not flight traits and power metabolism patterns have been density-dependent we carried out bioassays on the isolation of gregarious locusts and crowding of solitary locusts. One-day-old grownup solitary and gregarious locusts have been crowded and remoted for 13 d, respectively. Typical solitary and gregarious locusts have been used as controls. The degrees of expression of key genes concerned in power metabolism (COX4, ATPsyn, and NDUFA) have been measured. Gene expressions of solitary locusts after crowding therapy decreased considerably in contrast with these of typical solitary locusts. In contrast, the identical genes have been elevated dramatically in the course of the isolation of gregarious locusts (Fig. 5A). In step with the gene-expression profiles, CS exercise and CO2 manufacturing price have been decreased in response to crowding therapy and elevated in response to isolation therapy (Fig. 5B and C). Thus, the traits of primary power metabolism in solitary and gregarious locusts have been decided by inhabitants density experiences.

Power metabolism and flight trait are reshaped by inhabitants density modifications. G and S characterize gregarious and solitary locusts, respectively. (A) Expression profiles of key genes associated to cardio power metabolism in response to the crowding and isolation therapy, respectively (n = 8, Pupil’s t take a look at). (B) Citrate synthase enzyme exercise measurement (n = 8, Pupil’s t take a look at). (C) CO2 manufacturing price measurement (n = 11 people, Pupil’s t take a look at). Isolation therapies have been normalized to typical gregarious locusts, and crowding therapies have been normalized to typical solitary locusts. (D) Flight efficiency comparability after isolation therapy (n ≥ 30 people, Mann–Whitney U take a look at). Every dot represents a single particular person. (E and F) GSH/GSSG and H2O2 content material measurement of remoted locusts throughout flight (n ≥ 6, Pupil’s t take a look at). (G) Flight efficiency comparability after crowding therapy (n ≥ 30 people, Mann–Whitney U take a look at). (H and I) GSH/GSSG and H2O2 content material measurement of crowded locusts throughout flight (n ≥ 6, Pupil’s t take a look at). Time factors are proven as 0.25- and 1-h flight therapy, every time level is in comparison with relaxation situation samples. The info are proven as imply ± SEM, *P < 0.05, **P < 0.01, and n.s, not important.
The flight performances have been in contrast after isolation and crowding therapies, respectively. After isolation, the short-term flight and most flight speeds of gregarious locusts have been considerably elevated, however the flight distance and flight period have been decreased in 0.25- and 1-h flight assessments, respectively (Fig. 5D). In contrast, the crowding of solitary locusts considerably elevated their flight distance and flight period, however their common and most flight velocity have been decreased (Fig. 5G). Moreover, flight-induced oxidative stress was examined in response to crowding and isolation therapies. The isolation of gregarious locusts considerably elevated the technology of oxidative stress, which concerned the discount of GSH/GSSG and the elevation of H2O2 degree after flight therapy (Fig. 5 E and F). In contrast, the crowding of solitary locusts repressed their H2O2 degree and elevated their GSH/GSSG degree after flight therapy (Fig. 5 H and I). Due to this fact, the completely different flight traits of solitary and gregarious locusts have been decided by primary power metabolic profiles and ROS technology and will be reversed by altering the inhabitants density.
Dialogue
Our examine confirmed that the flight trait differentiation between solitary and gregarious locusts was primarily attributed to the divergence of power metabolism, which was decided by completely different experiences of inhabitants density. Sturdy power metabolism in flight muscle groups of solitary locusts facilitated their short-term flight skill and velocity. Nevertheless, their extended flight efficiency was restricted due to the overload in oxidative stress. In contrast, gregarious locusts exhibited reasonable cardio exercise of their flight muscle groups, can maintain flight at a comparatively low velocity, and preserve steady redox homeostasis (Fig. 6).

Mechanism of density-dependent flight trait variation of migratory locusts. At low inhabitants density, solitary locusts exhibit strong power metabolism exercise, which produces extra energy for short-term flight. Nevertheless, an overload of oxidative stress, which is a by-product of cardio exercise, inhibits their flight sustainability. In contrast, reasonable metabolic exercise in gregarious locusts maintains a comparatively decrease ROS technology, thereby facilitating the sustainability of their long-term flight.
Atmosphere-dependent flight capability polyphenism is an adaptive technique that underpins the ecological success of many bugs. This examine reported an attention-grabbing flight technique for the migratory locust to deal with modifications in inhabitants density. The migratory locust can remodel from a solitary type to a gregarious one at elevated inhabitants density (6). Solitary and gregarious locusts displayed completely different flight traits, and such differentiation completely matched the organic traits of the locust part. In case of meals shortage and absence, long-distance swarm migration happens in response to unfavorable situations (31, 32). Thus, the sustained flight skill of gregarious locusts is vital to looking for new meals assets or preferrred reproductive areas. In contrast, underneath low-density situations with ample meals provide, a brief dash skill is enough for solitary locusts to keep away from predators or to seek for mates. Due to this fact, along with a transition in physique shade and poisonous alerts (17, 33), flight trait variation is a crucial characteristic of locusts. Moreover, crowded situations reportedly enhance the flight efficiency of wing-monomorphic bugs, akin to African armyworm, beet webworm, and Oriental armyworm (34⇓–36). Thus, the flight trait variation of bugs could also be a widespread adaptive technique for a lot of wing-monomorphic bugs.
Gregarious and solitary locusts exhibited differential power metabolism of their flight muscle groups. Solitary locusts exhibited greater mitochondrial energetic storage (Acetyl-CoA and NADH), power metabolic gene-expression ranges, and metabolic enzyme actions of their flight muscle groups than the gregarious ones. After we repressed the metabolic exercise of locusts by flattening CS expression, their short-term flight velocity additionally decreased. Thus, the excessive ranges of power metabolism within the flight muscle groups of solitary locusts allowed high-speed and short-term flight. Flying animals primarily depend on the oxidation of carbohydrates and lipids to gasoline their flight (30, 37, 38). In locusts, carbohydrates are usually used on the preliminary stage of flight or throughout short-term flight. Conversely, long-term flight requires lipid oxidation (39). The oxidation of carbohydrates within the mitochondria can promote the speedy manufacturing of ATP, which may help the excessive power necessities of short-term dash flight (40). Nevertheless, lipid oxidation can help a considerably decrease most price of ATP turnover, indicating that the method can energy solely the decrease energetic necessities of long-term or migratory flight. Thus, on the preliminary stage of flight or throughout a short-term flight, the flight velocity is decided by carbohydrate consumptive price and mitochondrial actions.
The decrease ROS technology of gregarious locusts facilitates their skill to carry out a long-distance flight. In solitary locusts, considerably decreased uric acid and GSH ranges and elevated H2O2 ranges throughout flight indicated speedy ROS accumulation, which considerably repressed their flight sustainability. Oxidative stress is presumably a key determinant of restricted extended flight skill of locusts. Insect flight is a totally cardio course of. Cardio respiration within the mitochondria is the primary supply of ROS, and excessive cardio efficiency is all the time related to intense ROS technology (41). An overload in ROS manufacturing induces oxidative stress and harms cells or tissues by activating a series response of lipid peroxidation. Thus, sustaining a steady redox stability is necessary for flying adaptation (42). In our examine, gregarious locusts throughout flight maintained a redox stability via low-energy metabolism exercise and excessive plasmalogen content material. Average metabolic exercise can cut back ROS manufacturing price, and a excessive plasmalogen content material can present different phospholipid and lipoprotein particles safety in opposition to oxidative harm (43).
The connection between power metabolism exercise and the long-distance flight skill of animals is ambiguous. A number of metabolic patterns have been noticed in flying animals, particularly in bugs with long-distance flight efficiency. For instance, populations of the cotton bollworm Helicoverpa armigera, which has nice long-distance flight skill, have excessive lipid metabolic gene expression ranges (24). In contrast, the flight MR is decrease in migratory populations of the monarch butterfly than in nonmigratory populations (22, 24). Sturdy MR and excessive metabolic gene expression degree hardly present any optimistic contribution to the long-distance flight of locusts due to extreme oxidative stress accumulation. Thus, a stability between power provide and oxidative stress is necessary for the long-distance flight of locusts.
Ample power storage can be necessary for long-distance flights. Lipids are the most important power assets for extended flight and are primarily saved in fats our bodies in migratory locusts. In flight, saved TAGs are catalyzed into 1,2-diacylglycerol and transported via the hemolymph into the flight muscle groups (19). Gregarious locusts have greater TAG storage in fats our bodies, whereas solitary ones have a better AKH content material in corpora cardiac however have restricted extended flight efficiency (44). This phenomenon is defined by the solitary locusts have much less intense hyperlipemia response to flight in hemolymph and require extra AKH content material to mobilize power metabolism, which focuses on single prereproductive migratory flight (45, 46). In our examine, when solitary locusts have been compelled to fly continually for 1 h, which was for much longer than their spasmodic bouts of flight (Fig. 1F), no apparent discount in TAG content material was noticed in fats our bodies. Thus, we suggest that the brief flight period of solitary locusts isn’t restricted by comparatively low TAG storage in fats our bodies. Nevertheless, oxidative stress, which is a by-product of the upper metabolic profile, restricts the extended flight exercise of solitary locusts. Much less ROS technology and excessive TAG storage contributes to the long-distance flight of gregarious locusts.
Inhabitants density modifications can reverse the metabolic profiles in migratory locusts. Our examine indicated that locusts can change their flight traits and power metabolism patterns in response to the alteration in inhabitants density. Such speedy plasticity explains why locusts possess completely different phase-related flight traits inside one technology. Earlier research discovered that gregarious nymphs are extremely lively in habits and possess a better CO2 manufacturing price than solitary nymphs (16, 47). Right here, a totally reversed metabolic profile was noticed within the flight muscle groups of grownup locusts. These variations in resting MRs between nymphs and adults could also be associated to elements akin to developmental stage or habits. The density-dependent alterations in metabolism are systematic and presumably regulated by a number of physiological alerts and pathways. Due to this fact, additional research are wanted to elucidate the underlying molecular mechanisms.
As probably the most broadly distributed and some of the infamous locust species, the migratory locust, which has long-distance flight skill, stays a critical menace to meals safety. The locust adopted a decreased power metabolism price technique to accumulate much less ROS technology for long-distance flight. This discovering enhanced our understanding of the connection between power metabolism price and long-distance flight capability and gives worthwhile insights into the flight adaptation methods of flying animals.
Supplies and Strategies
Bugs and Experimental Therapies.
Locusts used on this examine have been from gregarious and solitary locust colonies maintained within the Institute of Zoology, Chinese language Academy of Sciences. Gregarious locusts have been cultured in giant bins (40 × 40 × 40 cm3) at a density of ∼300 bugs per container for no less than eight generations. Solitary locusts have been cultured alone in white metallic bins (10 × 10 × 25 cm3) equipped with recent air for no less than 10 generations earlier than experiment motion. This colony was maintained underneath a 14-h mild:10-h darkish photocycle regime at 30 ± 2 °C and ate up recent wheat seedlings.
One-day-old adults have been used for isolation and crowding therapies. For the isolation therapy, gregarious adults that eclosed inside 24 h have been reared underneath the solitary situation as described above. For the crowding therapy, the solitary adults that eclosed inside 24 h have been reared along with grownup gregarious locusts in giant bins. The remoted or crowded locusts have been dissected or used for the flight efficiency assessments on the 14th day put up eclosion.
Tethered Flight Take a look at and Flight Therapy.
Flight experiments have been performed on the identical time by utilizing computerized tethered flight mills housed on the Institute of Zoology, Chinese language Academy of Sciences. In a typical process, the flight arm (radius = 12 cm; flight circumference = 75 cm) was product of onerous plastic with the vertical axis secured between two magnets. The magnets ensured the negligible friction between the flight arm and corbelled vertical axis. Particular person locusts have been hooked up by the tergal aspect of the mesothorax to the top of the flight-mill arm by utilizing a skinny copper wire that fitted right into a sleeve suspended from the flight-arm terminal. This setup enabled the insect to fly rotationally in a horizontal aircraft. An infrared radiation sensor mounted beneath the flight arm detected a rotational variety of turns in order that the flight parameters have been recorded. The locust flight was induced by a fan positioned above the flight mill (if the interval of 30 s no sign is detected, blow for two s, at 1.5 m/s wind velocity). Within the flight traits take a look at, velocities have been calculated by dividing distance flown in a flight bout by the period of that flight bout after which averaged throughout all flight bouts throughout the take a look at interval. The flight distance was calculated by multiplying the flight arm circumference by the variety of laps flown throughout the take a look at interval. The recording of flight parameters and associated calculations are carried out routinely by a pc. The flight-mill system was positioned in a room underneath a photoperiod of 14 h mild:10 h darkish at 28 °C and 5% humidity.
Based mostly on the mass and flight information, we estimated the flight kinetic power (KE) throughout flight course of as KE = 1/2 × mass × velocity2 (joules) (28). Flight muscle mass was decided not directly by slicing the thorax (shorn of legs) and eradicating hint nonmuscular options akin to fat and elements of the digestive tract. Then, the thorax was weighed and soaked in 0.5 mol/L NaOH for twenty-four h. The cuticular residue was weighed after digestion of the muscle tissue (48).
Flight therapy, as distinguished from tethered flight traits take a look at, was used to research the differential regulatory mechanisms between gregarious and solitary locusts. For the pattern preparation of flight therapy, 14-d-old grownup male solitary locusts have been compelled to sustained flight for the full period of 0.25 and 1 h, respectively. Gregarious locusts have been handled in the identical method. Locusts that stopped flying have been artificially stimulated to proceed flight. The flight information generated by the flight therapy weren’t included within the evaluation of the locust flight traits detection. Flight muscle or fats physique was collected instantly after flight therapy and positioned in liquid nitrogen for additional assessments.
Physique Weight and Wing Measurement.
Physique weight was decided by weighing 14-d-old grownup male locusts on an digital stability. Wing size and width have been measured. The forewing and hindwing of every locust have been rigorously eliminated on the wing base by utilizing forceps and scissors. Utilizing a digital caliper, the forewing size, forewing width, hindwing size, and hindwing width have been measured. The wing landmarks used for every measurement have been as follows. Forewing size was from the bottom of the costal vein to the tip of the apical angle (roughly the apical finish of the fourth radial department). Forewing width was from the apical finish of the third radial department to the anal margin (a line roughly parallel to the physique). Hindwing size was from the bottom of the cubital vein to the apical finish of the third medial department, and hindwing width was from the apical finish of the subcostal vein to the anal finish of the third anal department.
RNA Sequencing and Information Processing.
Whole RNA was extracted from flight muscle (no less than three samples with six people per pattern) by utilizing TRIzol reagent (Invitrogen). Complementary DNA libraries have been generated utilizing NEBNext UltraTM RNA Library Prep Package for Illumina (NEB) following the producer’s suggestions, and index codes have been added to attribute sequences to every pattern. Uncooked information have been filtered, and the cleaned information have been mapped to the locust genome sequence with HISAT2 software program. DEGs have been analyzed by utilizing EdgeR software program. Moreover, unsupervised hierarchical clustering was carried out utilizing Clustal 3.0. RNA-sequencing information have been deposited within the Sequence Learn Archive Database of NCBI (BioProject PRJNA751147).
qRT-PCR.
The relative expression of messenger RNA (mRNA) was quantified with SYBR Inexperienced equipment (Roche) and LightCycler 480 instrument (Roche). The relative expression ranges of particular genes have been calculated and quantified utilizing the two −ΔΔCt methodology. Rp49 was thought of as endogenous management for mRNAs. Dissociation curves have been decided for every gene to substantiate distinctive amplification. The qPCR primers are listed in SI Appendix, Table S2.
Respiratory MR Measurement.
Respiratory MR assays have been carried out as described beforehand (49). The respiratory MR of the locusts was measured at 28 °C environmental temperatures by utilizing a respirometry system (Sable Methods Worldwide). We used respiratory fuel trade by estimating CO2 manufacturing price as a proxy for MR. The MR of the locust was examined in a closed-circuit system with a quantity of 116.7 mL. Every locust was enclosed inside a chamber of the circuit that had been positioned in an incubator. We now have a water-scrubbed system, containing calcium sulfate, to take away the water generated in the course of the measurement. The chamber, water-scrubbed system, and CO2 analyzer have been included within the closed circuit. Firstly of measurement, the circuit system was open to the air scrubbed of water and CO2 via a tube at a circulation price of 240 mL/min to stabilize the baseline. The circuit system was then transferred to a closed cycle, and the speed of CO2 manufacturing (VCO2) within the closed-circuit system was constantly recorded each second for no less than 5 min. The output of the CO2 analyzer was digitized (as soon as per second) and recorded utilizing the ExpeData information evaluation software program (Sable Methods). For the calculation of MR, we chosen the interval when locust respiration was steady and estimated the MR by calculating the change of CO2 quantity per unit time. MR was calculated because the CO2 produced per gram physique mass per minute utilizing the equation MR = VCO2 × quantity/physique mass, the place VCO2 × quantity is the change in quantity of CO2 in the course of the chosen take a look at time. The VCO2 is obtained from Expedata software program information and expressed as a decimal fraction. The amount was 116.7 mL of the closed circuit. All measurements have been performed from 10 AM to six PM, to attenuate the impact of circadian rhythms on the locusts.
RNA Interference.
The dsRNA of GFP and CS have been ready utilizing a T7 RiboMAX Specific RNAi system (Promega) following the producer’s protocol. About 3 µg⋅µL−1 dsRNA (complete = 9 µg) was injected into 11-d-old grownup locusts on the second ventral phase of the stomach. The dsGFP-injected group served because the management. After 72 h, the locusts have been dissected or used for the flight efficiency assessments. The primers for gene dsRNA synthesis are listed in SI Appendix, Table S2.
Enzyme Exercise Measurement.
CS exercise was measured utilizing a Citrate Synthase Exercise Colorimetric Assay Package (Biovision, K318-100) following the producer’s protocols. About 10 mg of tissues was homogenized on ice with 100 µL of ice-cold CS assay buffer and centrifuged at 10,000 × g for five min. Twenty microliters of supernatant was collected and added right into a 96-well plate, and the amount was adjusted to 50 µL with CS buffer. Then, 50 µL of response combine was added into every effectively containing samples, and absorbance (optical density at 412 nm) was measured instantly in kinetic mode at 25 °C for 20 to 40 min. The measured values have been normalized to protein ranges.
PYK enzyme exercise was measured by colorimetric assay utilizing an exercise Assay Package (Solarbio, BC0540) following the producer’s protocols. Then, 20 mg of flight muscle tissue of locust was totally homogenized with 400 µL ice-cold extraction buffer. After centrifuging the homogenized tissue at 8,000 × g at 4 °C for 10 min, the supernatant was collected and transferred right into a clear tube and saved on ice. The supernatant (20 µL) and response buffer (200 µL) have been used for the enzyme exercise assay. Absorbance was measured at optical density at 340 nm. HK exercise was measured with a Hexokinase Assay Package (Solarbio, BC0745). About 20 mg flight muscle tissue was homogenized with 400 µL ice-cold assay buffer and centrifuged for 10 min at 4 °C at 8,000 × g. Then, 200 µL response buffer was added into 10 µL supernatant, and absorbance at optical density at 340 nm was instantly measured in kinetic mode at 25 °C for five min. The measured values have been normalized to protein ranges.
Acetyl-CoA and NADH Measurement.
Acetyl-CoA ranges have been decided utilizing an Acetyl-CoA Assay Package (Sigma, MAK039-1KT). Contemporary muscle tissues have been homogenized with 1.0 M perchloric acid. The samples have been centrifuged at 10,000 × g for 10 min to take away insoluble materials. After focus, the supernatant was neutralized with 3 M potassium bicarbonate resolution. Resolution pH was maintained throughout the vary of 6 to eight. Acetyl-CoA ranges have been measured utilizing a premade response combination following the producer’s protocol after which normalized to protein ranges.
For NADH measurement, recent muscle tissue was frozen in liquid nitrogen, homogenized with 400 µL of NADH/NAD extraction buffer, and centrifuged for 10 min at 4 °C at 10,000 × g. Then, the supernatant was centrifuged for 20 min at 4 °C at 10,000 × g by utilizing a 10-kDa spin column (Abcam, ab93349). The filtrate was collected and used for NADH detection with a NAD/NADH Assay Package (Abcam, ab65348). The measured values have been normalized to protein ranges.
LD Staining.
LDs have been visualized by staining fats physique tissue with Nile pink. Fats physique tissues have been dissected and washed 3 times with 1× phosphate-buffered saline (PBS) and incubated for two h in Nile pink mounting medium (20% [vol/vol] glycerol in PBS, with a 1:10,000 dilution of 10% [vol/vol] Nile pink in dimethyl sulfoxide) at room temperature. After two or three washings with 1× PBS, the tissue was incubated for 10 min in Hoechst with 1:500 dilution in PBS after which washed twice with 1× PBS. All photographs have been taken utilizing a Zeiss LSM 710 confocal microscope at 40× magnification with an excitation wavelength of 543 nm and an emission wavelength of 626 nm. The dimensions of LDs was measured utilizing Zeiss ZEN software program.
TAG and Glycogen Measurement.
Whole TAG content material was estimated in keeping with earlier research (50). At the least three particular person tissues have been blended and 0.02 g of pattern was homogenized in 100 µL PBS containing 0.5% Tween-20 and incubated at 70 °C for five min. Then, all samples have been incubated with triglyceride reagent (Sigma, T2449) at 37 °C. After 30 min, the collected supernatant was transferred into 96-well plates after centrifugation and incubated with free glycerol reagent (Sigma, F6428) for five min at 37 °C and assayed utilizing SpectraMax Plus384 with a wavelength of 540 nm. TAG quantities have been decided by subtracting the quantity of free glycerol within the PBS-treated pattern from the full glycerol current within the pattern handled with triglyceride reagent. TAG ranges have been normalized to protein ranges in every homogenate via BCA assay (Thermo, 23228/23224).
Whole glycogen content material was decided utilizing a Glycogen Colorimetric/Fluorometric Assay Package (Biovision, K646–100) in keeping with the producer’s protocols. In a typical process, 10 mg of muscle tissue was homogenized with 300 µL of double-distilled H2O and boiled for 10 min. Then, the samples have been centrifuged at 14,000 × g for 15 min. The supernatant was reacted with 2 µL of hydrolysis enzyme combine for 30 min at room temperature. Lastly, we added 50 µL of response combine to every effectively for 30 min. The measured values have been normalized to protein ranges in every homogenate.
Metabolomic Profiling.
The metabolomic profiling evaluation included pattern preparation and extraction, metabolite detection, metabolomic information preprocessing, chromatographic peak alignment, and statistical evaluation. Particulars are included in SI Appendix.
Paraquat Injection.
For paraquat therapy, 12-d-old male adults have been chosen randomly and separated into 4 teams (no less than 20 people per group). Amongst them, the saline-injected (2 µL per locust) group served because the management for oxidative harm. The remaining three teams have been injected with 2 µL of paraquat (Macklin, P814066) at concentrations of 4, 20, and 100 mmol⋅L−1. At 48 h posttreatment, all handled locusts have been dissected or used for flight efficiency assessments. The dissected pattern tissues have been maintained in liquid nitrogen.
Oxidative Stress Measurement.
A Hydrogen Peroxide Assay Package (Beyotime, S0038) was used to find out H2O2 content material. Flight muscle tissues have been harvested and washed with PBS. Ten milligrams of tissue was resuspended in 200 µL of extraction lysis buffer. After homogenization with a vortex shaker, the lysate was centrifuged at 12,000 × g in a microcentrifuge for five min at 4 °C. A 50-µL aliquot of supernatant was used for H2O2 content material dedication with 100 µL of detection buffer. Blended samples have been incubated for 30 min at room temperature and assayed utilizing SpectraMax Plus384 with a wavelength of 560 nm. Ratios of decreased to oxidized glutathione (GSH/GSSG) have been measured utilizing a GSH/GSSG Ratio Detection Assay Package (Beyotime, S0053). Ten milligrams of flight muscle tissue was homogenized with 100 µL of assay buffer and centrifuged for 10 min at 4 °C at 10,000 × g. First, complete glutathione content material was measured. Ten-microliter supernatant samples and 150 µL response combine have been incubated at room temperature for five min. Then 50 µL of the 0.5 mg/mL NADPH resolution was added and blended effectively. The absorbance at 412 nm was instantly measured in kinetic mode at 25 °C for 25 min. Second, one other pattern was used for GSSG content material measurement. A 50-µL supernatant pattern was reacted with 2.5 µL GSH scavenger for 60 min. Then, 10 µL of the answer from the earlier step and 150 µL response combine have been incubated at room temperature for five min. Then, 50 µL of the 0.5 mg/mL NADPH resolution was added and blended effectively. The absorbance at 412 nm was instantly measured in kinetic mode at 25 °C for 25 min. Lastly, the quantity of GSH is the same as the full glutathione minus GSSG.
Information Availability
RNA-sequencing information have been deposited within the Sequence Learn Archive Database of NCBI (BioProject PRJNA751147). All different examine information are included within the article and/or SI Appendix.
Acknowledgments
We gratefully acknowledge Jia Yu for the locust pattern assortment. We additionally thank Weiguo Du for useful feedback. This examine was supported by the Nationwide Pure Science Basis of China (grants 32088102 and 31920103004).
Footnotes
- Accepted November 10, 2021.
-
Writer contributions: B.D. and L.Ok. designed analysis; B.D., W.G., and L.Ok. carried out analysis; B.D., D.D., C.M., W.G., and L.Ok. analyzed information; and B.D., D.D., and L.Ok. wrote the paper.
-
Reviewers: D.D., The Ohio State College; and J.H., Arizona State College.
-
The authors declare no competing curiosity.
-
See online for associated content material akin to Commentaries.
-
This text incorporates supporting data on-line at https://www.pnas.org/lookup/suppl/doi:10.1073/pnas.2115753118/-/DCSupplemental.
- Copyright © 2021 the Writer(s). Printed by PNAS.












{kind=link}