
Lipid membranes are self-assembled viscoelastic bilayers separating cells and their organelles from their atmosphere (1). They’re straightforward to bend however immune to stretching: Their lysis pressure—the stress at which they break—is excessive, within the vary of some millinewtons per meter (2). This excessive worth protects cells towards lysis upon processes that stretch the cell membrane, but it surely nonetheless can not defend from the large stretch generated by hypotonic shocks (3). Plasma membrane pressure arises from the mixed contributions of osmotic strain, in-plane pressure, and cytoskeletal forces (4). The cytoskeleton is intimately linked to all processes regulating membrane pressure, specifically cell quantity regulation (5). For instance, hypotonic shocks not solely are answerable for rising membrane pressure (6, 7) but in addition induce the degradation of vimentin (8) and a reorganization of actin filaments (9, 10), with out strongly affecting microtubules (8). The cytoskeleton regulates membrane pressure by setting its worth by means of energetic pressure era and by establishing a membrane reservoir that buffers acute modifications in pressure (11). This membrane reservoir is saved round protruding actin-based constructions resembling ruffles, filopodia, and microvilli.
A part of this membrane reservoir can also be saved in caveolae, that are plasma membrane invaginations fashioned by the meeting of Cavin1-3 and Caveolin1-2, and their disassembly buffers membrane pressure enhance (6). Cell quantity regulation throughout osmotic modifications includes mechanosensitive ion transporters and channels (12, 13) regulated by membrane pressure (14). How their exercise is coupled to the cytoskeleton is underneath debate (15). They comprise volume-regulated anion channels (VRACs), sodium–hydrogen antiporters (NHEs), and Na-Okay-Cl cotransporters (NKCC1). VRACs are activated by hypotonic stress (16) and are distinctive in transporting small natural osmolytes—specifically, taurine—along with anions (17). NHEs inhibition prevents regulatory quantity enhance of cells (18). Cells have advanced to answer modifications in membrane pressure and management its affect on many processes important to cell life (19). The genetic response to an osmotic stress has been studied extensively. This pathway partly consists of activating genes concerned within the synthesis or degradation of osmo-protectant molecules (resembling glycerol in yeast and amino acids in mammalian cells) and their subsequent secretion (20, 21). Nevertheless, the timescale of the genetic response to osmotic stress—on the order of tens of minutes—can not account for the cell’s fast resistance to stretch (18, 21). The grasp regulator of plasma membrane pressure might be Goal of Rapamycin Advanced 2 (TORC2) (22) and its mammalian homolog mTORC2. TORC2 and mTORC2 are assembled across the kinase TOR (mTOR), which can also be a part of TORC1 and mTORC1. Whereas TORC1 and mTORC1 are concerned in cell quantity regulation (23, 24), TORC2 and mTORC2 are concerned in membrane pressure regulation (7, 25, 26). Certainly, TORC2 signaling will increase instantaneously upon membrane pressure enhance (27) in addition to mTORC2 exercise (7) and reduces upon pressure loss (25). TORC2 regulates endocytosis by means of membrane pressure (26), but in addition actin polymerization, as knockdown of mTORC2, however not mTORC1, prevents actin polymerization (28) and will increase membrane pressure (7). The elevated pressure might be linked to the position of cortical actin in folding the membrane into ruffles, controlling the dimensions of the membrane space buffer (29).
Regardless of its simple significance, the mechanisms driving the regulation of membrane pressure throughout osmotic shocks in relation to cell quantity modifications are nonetheless poorly understood. Qualitatively, membrane pressure has been reported to lower in response to hypertonic shocks (30, 31), whereas research have reported that it both stays fixed (31, 32) or will increase (6, 30, 33) upon hypotonic shock. The relation between osmolarity and cell quantity change is captured by the Ponder/Boyle/Vant’Hoff (PBVH) relation whereby cell quantity modifications till the osmotic strain of its contents matches that of the extracellular medium (34). This relation includes an osmotically inactive quantity (V OI), which represents the minimal quantity occupied by tightly packed mobile constituents at very excessive hypertonicity (23, 35, 36). Nevertheless, whereas the PBVH relation describes the modifications in cell quantity in response to an osmotic shock, the membrane pressure response to such shocks has by no means been quantitatively studied. On this examine, we elucidate quantitatively the coupling between cell quantity and membrane pressure in single cells throughout osmotic shocks utilizing time-resolved methods.
Outcomes
Cell Quantity Change upon Osmotic Shock
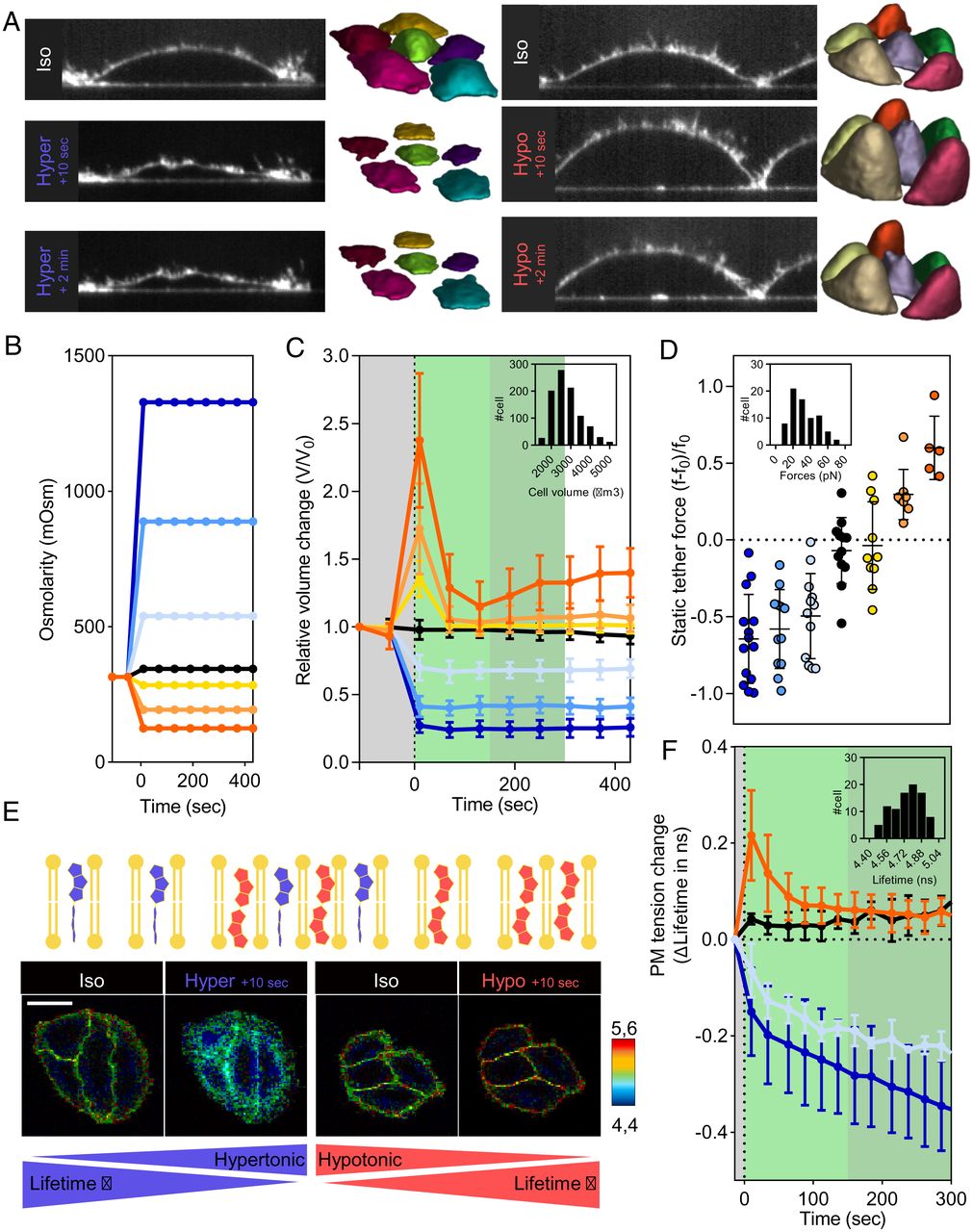
We measured quantity modifications in HeLa Kyoto cells labeled with the plasma membrane probe CellMask and uncovered to stepwise osmotic shocks (Fig. 1 A and B and Supplies and Strategies). Volumes had been obtained by segmenting time-lapse three-dimensional (3D) confocal stacks of CellMask photos. Imaging large unilamellar vesicles (GUVs), we estimated the error of our quantity quantification to be lower than 5%, in any case required corrections (SI Appendix, Fig. S1A). The typical cell quantity peaked at roughly 2.4 occasions the preliminary quantity, a couple of seconds after a hypotonic shock (120 mOsm). It subsequently recovered, however solely partially, leaving a 15% quantity enhance after 10 min (Fig. 1C). Weaker dilutions (50% and 25%) led to decrease peaks (Fig. 1C). Cell porosity measured utilizing propidium iodide (PI) in hypotonic medium was null (SI Appendix, Fig. S1B). Conversely, hypertonic shocks led to a fast quantity lower inside seconds, adopted by a 10-min plateau. Cells weren’t porous (SI Appendix, Fig. S1B) and cell metabolism after 1 h in hypertonic buffer as much as 900 mOsm was greater than 60% (SI Appendix, Fig. S1C) and equal to 100% if cells had been left to get better in a single day in isotonic medium (SI Appendix, Fig. S1D). In essentially the most excessive hypertonic situations (3,500 mOsm), cell quantity decreased by as much as 90% (SI Appendix, Fig. S1 E and F). Earlier than the osmotic shocks, cell quantity distributions had been broad (Fig. 1C) whereas relative cell quantity modifications had been extremely reproducible. Utilizing an EGFP-LaminB1 secure cell line, we measured nuclei and cell quantity modifications and confirmed that cell quantity modifications had been basically as a consequence of cytoplasmic quantity modifications (SI Appendix, Fig. S2). To evaluate the robustness of the restoration dynamics, we carried out cell quantity measurements utilizing a distinct cell kind (HL-60/S4) utilizing real-time deformability cytometry (RT-DC)—a high-throughput approach permitting for fast characterization of 1000’s of cells (37) (SI Appendix, Fig. S3A). After making use of the osmotic shocks to HL-60/S4 cells earlier than loading them into RT-DC, we confirmed our earlier commentary: Cell quantity in a hypotonic medium peaked and recovered whereas cell quantity in a hypertonic medium abruptly and stably decreased (SI Appendix, Fig. S3 B and C). Nevertheless, the attribute time of the restoration might be cell kind dependent and whether or not cells are adherent or not might affect this attribute time (38). Our outcomes present that the amount modifications related to osmotic shocks are fast, with an uneven restoration, fast for hypotonic shocks and really gradual for hypertonic shocks. Based mostly on these observations and former outcomes (25, 27), we hypothesize that the regulation of cell response to hypotonic shocks is distinct from the regulation of cell response to hypertonic shocks.

Osmotic shocks have an effect on cell quantity and membrane pressure. (A) Consultant facet view of cell with CellMask staining and corresponding 3D reconstruction of cell quantity utilizing Limeseg (Left, hyperosmotic; Proper, hypoosmotic). (B) Osmolarities (mOsm) of cell media with time for the completely different stepwise osmotic situations. Osmotic situations correspond to 120 mOsm in darkish orange, 190 mOsm in gentle orange, 285 mOsm in yellow, 315 mOsm in black, 540 mOsm in gentle blue, 890 mOsm in medium blue, and 1,330 mOsm in darkish blue. Coloration code is maintained. (C) Averaged cell quantity dynamics underneath osmotic shock (grey, earlier than shocks; gentle inexperienced, short-term response; darkish inexperienced, long-term response). Statistics are as follows: darkish orange, R > 3, n = 28; gentle orange, R > 3, n = 20; yellow, R > 3, n = 40; black, R > 3, n = 66; gentle blue, R > 3, n = 62; medium blue, R > 3, n = 47; and darkish blue, R > 3, n = 43. Inset exhibits cell quantity distribution in isotonic medium earlier than osmotic shocks (R > 50, n = 959). (D) Relative change of tether pressure instantly after osmotic shocks (averaged over 10 s) for various osmotic shocks. Inset exhibits tether pressure distribution in isotonic medium earlier than osmotic shocks (R = n =73). (E) Fluorescence lifetime of the Flipper-TR probe reviews membrane pressure modifications. Proven are FLIM photos of Flipper-TR lifetime values (colorscale) of cells subjected to osmotic shocks. (F) Dynamics of the change of pressure as measured by Flipper-TR lifetime (grey, earlier than shock; gentle inexperienced, short-term response; darkish inexperienced, long-term response). Osmotic situations corresponds to 120 mOsm in darkish orange (R = 7, n > 28), 315 mOsm in black (R = 3, n > 12), 600 mOsm in gentle blue (R = 3, n > 12), and 1,330 mOsm in darkish blue (R = 5, n > 20). Inset exhibits distribution of the cell common Flipper-TR lifetimes in isotonic medium earlier than osmotic shocks (R > 100, n > 400).
Membrane Pressure Adjustments Are Coupled to Cell Quantity Adjustments
We then measured modifications of membrane pressure after osmotic shocks. When membrane tubes are pulled from the cell membrane utilizing beads held with optical tweezers, the pressure required to statically maintain the tube is a direct measurement of membrane pressure. The distribution of tube forces earlier than osmotic shock was broad: 27 ± 18 pN (Fig. 1D). Adjustments within the tube pressure upon osmotic shocks had been quick (a couple of seconds) and beneath the temporal decision of the cell quantity acquisition (SI Appendix, Fig. S3 D and E). Curiously, modifications of the tube pressure averaged over 10 s instantly following the shock had been in line with the depth of the shocks (Fig. 1D and SI Appendix, Fig. S3 F and G). To comply with the dynamics of pressure in actual time, we used the molecular probe Flipper-TR (fluorescent lipid pressure reporter [FliptR]) that reviews modifications of membrane pressure by means of modifications of its fluorescence lifetime (Fig. 1E) (31, 39). Per tube pulling experiments, the lifetime distribution of Flipper-TR in cell membranes earlier than shock was broad: 4.76 ± 0.15 ns (Fig. 1F), and the lifetime modified inside seconds after shock (Fig. 1 E and F). We noticed an asymmetry in lifetime measurements throughout restoration much like the one noticed for quantity measurements: Membrane pressure peaked and recovered inside seconds after hypotonic shock whereas it decreased inside seconds after hypertonic shock and continued reducing at some stage in the experiment, though at a decrease charge (Fig. 1F). Our outcomes present that membrane pressure variations after osmotic shocks qualitatively comply with cell quantity modifications. To additional examine potential variations between quantity and pressure restoration, we studied them for two h after a gentle hypertonic shock (600 mOsm). Cell quantity slowly began recovering 15 min after the shock (SI Appendix, Fig. S4A). Per refs. 40 and 41, restoration was nonetheless incomplete 2 h after the shock. Membrane pressure had very completely different restoration dynamics: It began recovering solely 50 min after the shock, however the restoration is quick and full after 80 min (SI Appendix, Fig. S4B). These observations present that quantity and pressure responses after hypertonic shocks might be decoupled in the long run. This uncoupling could also be as a consequence of reactivation of energetic cell processes, resembling endocytosis or lipid metabolism, which can be triggered by a genetic response to the osmotic shock. Certainly, by reintroducing options with physiological osmolarities (315, 250, and 160 mOsm) on the time when quantity restoration had began, however not pressure restoration (20 min after shock; SI Appendix, Fig. S4 A and B), we noticed that quantity and pressure recoveries had the identical dynamics (SI Appendix, Fig. S4 C–F), however solely hypotonic medium (160 mOsm) led to a full restoration (SI Appendix, Fig. S4D). Altogether, these outcomes help the notion that the coupling between cell quantity and pressure responses to osmotic shocks is stored not less than 20 min after the shock.
Quantitative Coupling of Cell Quantity to Osmotic Shocks
To quantitatively seize the connection between the osmotic strain of the cell and its quantity (Fig. 2 A and B and SI Appendix, Fig. S5 A and B), we used the PBVH equation of state (34, 42)[1]the place P is the osmotic strain of the intracellular medium, V is the cell quantity, and P0 and V0 are values of P and V underneath isotonic situations. Eq. 1 assimilates the contents of the cell into an answer of particles with steric repulsions and in any other case negligible interactions, with the sum of the particles excluding volumes equal to V OI. The cell quantity thus can’t be compressed beneath the “osmotically inactive quantity” V OI. Eq. 1 is in glorious settlement with each our hypertonic and hypotonic knowledge (Fig. 2 A and B), when the amount is estimated on the time of hypotonic peak (10 s after the shock, proven in Fig. 1C and SI Appendix, Fig. S1F). Curiously, throughout and after the restoration section (>10 s after shock) quantity values diverged from this linear relation solely within the hypotonic situations, reflecting the asymmetry of the restoration (Fig. 2 A and B). A single-parameter match yields equal to about 10% of the preliminary cell quantity, smaller than earlier estimates (43). Utilizing RT-DC values on HL-60/S4 cells on the peak, we additionally discover a good settlement with , which is about 30% of its preliminary cell quantity. This distinction might be defined by the smaller vary of osmotic strain measured for HL-60/S4 (SI Appendix, Fig. S5B) or by the intrinsic overestimation of cell volumes in RT-DC. It is also that HL-60/S4 cells might also have a bigger osmotically inactive quantity, as it’s recognized to be cell kind particular (36).

Quantitative coupling of cell quantity and cell membrane pressure to osmotic shocks. (A) Normalized quantity modifications () as a perform of osmotic strain ratio () simply after osmotic shocks (stable circle) and eight min after the osmotic shock (open circle, restoration section) in comparison with the prediction of Eq. 1 (crimson line). Coloration code is maintained from Fig. 1. Statistics are (R > 3, n = 28) for darkish orange, (R > 3, n = 20) for gentle orange, (R > 3, n = 40) for yellow, (R > 3, n = 66) for black, (R > 3, n = 62) for gentle blue, (R > 3, n = 47) for medium blue, and (R > 3, n = 43) for darkish blue. (B) Normalized quantity modifications () as a perform of osmotic strain ratio () simply after osmotic shocks in comparison with the prediction of Eq. 1 (crimson line). Statistics are the identical as in A. (C) Refractive index photos of cells underneath osmotic shocks. (D) Protein focus modifications () in line with strain utilized (). Every level corresponds to at least one experiment containing not less than two cells. Evaluation is carried out on particular person cells. (E) Calculated dry mass of cells versus normalized strain (). Every level corresponds to at least one experiment containing not less than two cells. Evaluation is carried out on particular person cells. (F) Consultant FIB-SEM photos of the membrane tubulations in isotonic medium and hypertonic medium. (Scale bar: 2 µm.) (G) Scheme describing the parameters of the idea. (H) Relative modifications of membrane space () versus relative modifications of membrane pressure () in comparison with the prediction of Eq. 3 (crimson line). Coloration code is maintained from Fig. 1. Statistics are (R = n = 5 in x; R > 3, n = 28 in y) for darkish orange, (R = n = 8 in x; R > 3, n = 20 in y) for gentle orange, (R = n =10 in x; R > 3, n = 40 in y) for yellow, (R = n = 12 in x; R > 3, n = 66 in y) for black, (R = n = 12 in x; R > 3, n = 62 in y) for gentle blue, (R = n = 12 in x; R > 3, n = 47 in y) for medium blue, and (R = n = 14 in x; R > 3, n = 43 in y) for darkish blue. (I) Normalized pressure () versus normalized strain () and prediction obtained utilizing Eq. 4. Statistics are the identical as in F. (J) Scheme describing the idea.
Estimating the Osmotically Inactive Cell Quantity
Our verification of the PBVH relation yields two stunning outcomes: First, we discover that it holds for a really giant vary of osmotic pressures in HeLa cells, bigger than beforehand examined. Second, the V OI represents a smaller proportion of V O [only 10%, compared to values between 7 and 50% in other studies (36, 44, 45)]. Because the V OI is obtained from a linear match, the vary of information and the system used for the match can strongly change the V OI. We examined this chance by altering the vary of information used for the match and present that utilizing smaller ranges all the time supplies bigger V OI (Supplies and Strategies). One might argue that the bigger vary of osmotic shocks we used might artificially give decrease values of V OI. To have an unbiased estimate of the V OI, we sought to measure the protein mass of the cell and evaluate the hydrated minimal quantity of this mass to our V OI values. For this, we used optical diffraction tomography, a 3D tomographic label-free approach, to measure the cell’s refractive index (RI) (46), therefore giving a direct entry to modifications of cell mass and protein focus. Cells in isotonic situations had a median RI = 1.37 ± 0.01. A couple of seconds after a hypertonic shock (1,330 mOsm), cells had a rise in RI to 1.42v ± 0.01, whereas underneath a hypotonic shock (120 mOsm) the RI decreased to 1.35 ± 0.01 (Fig. 2C). RI will increase linearly with rising protein focus (47). In our experiment, the RI of single cells modified proportionally to the utilized osmotic strain (SI Appendix, Fig. S5C). This means that the protein focus modifications proportionally to the osmotic strain (Fig. 2D), which is totally in line with our discovering that cell quantity modifications proportionally to the osmotic strain (Fig. 2 A–C). Extracting the focus from the RI and understanding the common cell volumes permit for calculation of the dry mass for every osmotic situation (Fig. 2E). The typical dry mass of single HeLa cells was 305 ± 98 pg (SI Appendix, Fig. S5D) and, as anticipated, is fixed all through all osmotic shocks (Fig. 2E). To immediately measure modifications of focus for a single protein throughout the cytoplasm, we measured the relative change of fluorescence depth of cells overexpressing cytosolic GFP over time. It additionally diversified proportionally to the osmotic strain (SI Appendix, Fig. S5 E–G). Thus, no important quantities of intracellular proteins had been exchanged with the atmosphere, in settlement with earlier research (26, 33). Our measurement of the dry mass (Fig. 2E) additionally enabled an estimation of the V OI. Multiplying the common quantity of hydrated proteins per unit of dry mass (0.73 mL/g) (48) by the dry mass, we discovered , in settlement with our PBVH match.
Quantitative Coupling of Membrane Pressure to Quantity Adjustments and Osmotic Stress
Because the cell quantity modifications, so do the stress and space of its membrane (Fig. 1 D–F and SI Appendix, Fig. S3 D–G). To compute the relation between these portions, we reasoned that the cell membrane shouldn’t be completely flat, however is as an alternative partly folded into tubular protrusions known as filopodia, induced by the motion of polymerizing actin. Many different proteins take part within the formation of these membrane protrusions, specifically inverted BAR (I-BAR) area proteins (49). Whereas different constructions than filopodia resembling caveolae might comprise substantial quantities of floor, in our cells, the biggest a part of the membrane is saved in filopodia constructions as seen from centered ion beam–scanning electron microscopy (FIB-SEM) 3D photos (Fig. 2F). Protrusions in hypertonic medium (800 mOsm) are longer and bigger than protrusions in isotonic medium (Fig. 2F). To account for the ensuing discount in obvious membrane space, we use a easy mannequin impressed by ref. 50 (Fig. 2G). Within the mannequin, the membrane exists in both of two states, tubulated or flat (Fig. 2F), and is at equilibrium with a chemical reservoir of proteins supplied by the cell’s cytoplasm. When a protein binds to the membrane, one of many tubular protrusions grows. The tubulated membrane space thus will increase by a, and the untubulated space decreases by the identical quantity. Observe that the identical reasoning might be utilized to spherical buds. The free vitality of the system is lowered by the binding free vitality µ of the protein and elevated by σa equal to the quantity of labor that should be carried out towards the membrane pressure σ to scale back the untubulated space. The free vitality µ regroups the protein entropy loss upon binding and the protein interplay with the membrane in addition to the elastic value of curving a membrane space a right into a tube. As every tube comprises many proteins, we neglect their translational free vitality in addition to the energetic value of curving the membrane at their tip and base. Denoting the overall membrane space by A, we are able to thus signify the membrane by a set of unbiased patches of space a, every of which might be tubulated or untubulated. The free vitality distinction between the 2 states reads , implying that the partition perform of the membrane reads , the place the single-patch partition perform reads[2]the place okBT is the thermal vitality. Denoting the obvious membrane space by A, the tubulated space is given by , implying that[3]
Setting the isotonic pressure σ0 to its measured imply , the match of this prediction to the info (Fig. 2H) yields a membrane space per protein and a binding free vitality , comparable to a binding free vitality per unit space . This worth is in step with estimates obtained for a number of protein coats (51) and extra usually for energies of molecular interactions with surfaces (52). For these parameter values, we estimate that the ratio of complete membrane space to membrane space within the isotonic state is . To acquire an extra indication of the boldness interval over our fitted values, we estimate the magnitude of the modifications within the values of a and µ that result in a noticeably worse match of the experimental knowledge (which we formally outline as an enhance of the sum of sq. residuals). We discover that such modifications are of the order of 15% for a and 10% for µ. To narrate the change in membrane pressure with the utilized osmotic strain, we mix the membrane’s tension-strain relation Eq. 3 with the PBVH equation of state Eq. 1 underneath the belief that the cell undergoes an roughly homogeneous dilation (contraction) when subjected to a hypoosmotic (hyperosmotic) shock. The final assumption implies that the amount and space of the cell respectively scale just like the dice and the sq. of its typical lateral dimension. This means , which we mix with Eqs. 1 and 3 to yield[4]
Eq. 4 yields a prediction for the dependence of the membrane pressure on osmotic strain, which is in good settlement with our knowledge (Fig. 2I). These outcomes strongly help the notion that the short-term responses of cell quantity and membrane pressure are predominantly mechanical and thermodynamic and encompass a passive equalization of the internal and outer osmotic pressures accompanied by an unfolding of membrane ruffles (Fig. 2J). To additional examine the position of membrane ruffles in cell osmotic response, we first studied the position of the cytoskeleton within the cell response to osmotic shocks.
Actin and Microtubule Dynamics throughout Cell Osmotic Response
As in our system, many of the protrusions concerned in buffering space enlargement appeared to be filopodia-like constructions, we sought to check the position of cortical actin within the osmotic shock response. We first imaged the dynamics of the actin cortex throughout osmotic shocks utilizing SiR-Actin. Upon hypotonic shock, we noticed cell blebbing concomitant with cortical actin depolymerization (SI Appendix, Fig. S6A). Blebs then prolonged and merged into a big membrane dome (SI Appendix, Fig. S6A, facet view). By quantifying cortical actin fluorescence, we noticed an entire repolymerization of the cortex 4 min after the shock (SI Appendix, Fig. S6B), to a worth greater than the preliminary worth. Following a hypertonic shock, the actin cytoskeleton appeared extra condensed, and its fluorescence depth step by step elevated with time (SI Appendix, Fig. S6B). We additionally adopted the conduct of microtubules utilizing SiR-Tubulin. After hypotonic shocks, microtubules additionally depolymerized and appeared extra condensed after a hypertonic shock, however to a smaller extent than actin (SI Appendix, Fig. S6 C and D). These outcomes advised that actin is strongly depolymerized by the unfolding of the membrane upon hypotonic shocks however then repolymerizes to a thicker cortex when cell quantity has totally recovered.
Influence of Cytoskeleton Inhibition or Stabilization on Cell Osmotic Response
To check this speculation, we used latrunculin A to depolymerize the F-actin or jasplakinolide to stabilize it (Fig. 3A). We then adopted the cell quantity and pressure modifications with time and in contrast them to these in untreated cells. First, we noticed that dimethyl sulfoxide (DMSO), the solvent of inventory options for the medicine we used, lowered the height of quantity after hypotonic shocks with out affecting the height of pressure in management experiments, as a result of DMSO elevated the osmolarity of hypotonic options (Supplies and Strategies). As described beneath, not one of the medicine used affected the response to hypertonic shocks (Fig. 3B and SI Appendix, Fig. S7A), in line with the hypertonic response being basically passive. Equally, each medicine had little impact on the preliminary peak in cell quantity after hypotonic shock, in line with the short-term response to hypotonicity being passive and with a earlier report on nonadherent cells (38). Nevertheless, latrunculin radically modified the later-time restoration in comparison with that in nontreated and jasplakinolide-treated cells. Certainly, the amount of latrunculin-treated cells partially recovered after the preliminary peak, however then diverged a couple of minutes after shock (Fig. 3B). Against this, the amount of jasplakinolide-treated cells advanced equally to that of nontreated cells (Fig. 3B), though over a shorter timescale. Curiously, the stress dynamics of each latrunculin- and jasplakinolide-treated cells had been utterly decoupled from quantity dynamics, as no peak, and thus no restoration, was noticed (Fig. 3C). Depolymerizing actin led to smaller preliminary pressure (11) and smaller cell quantity (SI Appendix, Fig. S7B) inducing extra membrane reservoir accessible, which may clarify the absence of membrane pressure enhance upon hypotonic shock (Fig. 3C). Stabilization of the actin additionally led to no enhance of membrane pressure underneath hypotonic shocks; we hypothesized that actin stretches to counterbalance the stress as an alternative of stretching the membrane. Thus, the actin cortex is a serious regulator of the coupling between membrane pressure and quantity dynamics (Fig. 3C). Depolymerization of microtubules with nocodazole had restricted results on the amount dynamics after hypotonic shocks but in addition decoupled pressure from quantity modifications, as no pressure modifications had been noticed (Fig. 3 B and C). Conversely, stabilizing microtubules with taxol clearly affected the dynamics of quantity modifications, as its peak was considerably smaller than in nontreated cells, and no restoration was noticed. The smaller quantity enhance of taxol-treated cells has additionally been noticed in nonadherent cells (38) and might be as a result of greater preliminary quantity, which would cut back the dimensions of membrane space buffer (SI Appendix, Fig. S7B). Perhaps due to this decrease space buffer, taxol-treated cells have the next pressure (11) and a stiffer membrane (53), which can clarify why cell quantity can not get better after its additional reasonable enhance (Fig. 3B). However, nocodazole-treated cells are softer (53) and have a decrease preliminary quantity (SI Appendix, Fig. S7B), which might be related to a bigger membrane space buffer. Per this, and as seen for cells handled with latrunculin, pressure doesn’t change upon hypotonic shock in nocadazole-treated cells (Fig. 3C). These outcomes present that pressure dynamics might be decoupled from quantity dynamics when actin and microtubule turnover is affected. As seen for taxol-treated cells, it’s potential to qualitatively change the amount and pressure response to osmotic shock, whereas preserving their coupling. Lastly, not one of the therapies affected the hypertonic response, supporting additional that cells reply passively to this situation or not less than with out the involvement of the cytoskeleton (SI Appendix, Fig. S7C). Whereas these outcomes clearly present that the cytoskeleton is a necessary actor within the cell response to osmotic shocks, a number of questions remained. Since actin was depolymerized throughout many of the restoration after hypotonic shocks, how is that this restoration achieved? Additionally, the tight coupling between quantity and pressure responses means that it’s underneath the management of a signaling pathway. Particularly, we puzzled what might be the position of caveolae, described as membrane pressure buffering constructions (6) and mTOR, a grasp signaling kinase concerned in controlling membrane pressure and membrane folding (7, 24).

Cytoskeleton controls the long-term response of cells to osmotic shocks. (A) Illustrations of cytoskeletal drug results. (B) Single-cell quantity dynamics of cells handled with latrunculin A, jasplakinolide, nocodazole, or taxol throughout hypotonic shocks (120 mOsm, circle), isotonic situations (315 mOsm, sq.), and hypertonic shocks (700 mOsm; P/P0 = 2, triangle). Statistics are R > 3 for each experiment. For latrunculin A, n = 5 for management, n = 4 for hypo, n = 15 for hyper; for jasplakinolide, n = 6 for management, n = 8 for hypo, n = 10 for hyper; for nocodazole, n = 8 for management, n = 8 for hypo, n = 6 for hyper; for taxol, n = 10 for management, n = 19 for hypo, n = 12 for hyper. (C) Membrane pressure dynamics of cells handled with latrunculin A, jasplakinolide, nocodazole, or taxol throughout the identical shocks as in B. Statistics are n > 4 for each experiment. For latrunculin A, R = 2 for management, R = 6 for hypo, R = 4 for hyper; for jasplakinolide, R = 4 for management, R = 10 for hypo, R = 3 for hyper; for nocodazole, R = 2 for management, R = 6 for hypo, R = 4 for hyper; for taxol, R = 2 for management, R = 3 for hypo, R = 4 for hyper.
Influence of Cavin1-Knockout on Cell Osmotic Response
We examined the position of caveolae in cell osmotic response through the use of HeLa Cavin1-KO, through which we totally invalidated the expression of the important part of caveolae Cavin1 (Fig. 4A and SI Appendix, Method). In these cells, we noticed that maximal cell quantity enhance underneath hypotonic shock corresponds to , much like that for management cells transfected with CRISPR Cas-9 with out information RNA (known as CRISPR management within the following) (Fig. 4B and SI Appendix, Fig. S7D). However, membrane pressure enhance was considerably greater in Cavin1-KO cells than in CRISPR management cells (Fig. 4C and SI Appendix, Fig. S7E). Utilizing the calibration of lifetime to membrane pressure from HeLa cells (31), a distinction of = 0.2 ns in hypotonic shocks corresponds to a doubling of membrane pressure. The relative quantity and membrane pressure modifications underneath hypertonic shocks (600 mOsm) had been the identical between the CRISPR management and the Cavin1-KO and so they weren’t completely different from the management. We interpreted these outcomes as attributable to a lowered membrane reservoir in Cavin1-KO cells, hypotonic shocks would quickly unfold the membrane space buffer, limiting quantity enlargement and rising pressure massively. Our observations are in line with caveolae accounting for a restricted portion of pressure buffering (6).

Caveolae and mTORC2 are concerned in membrane pressure regulation. (A) Schematic of membrane ruffles interactions with caveolin1-2, cavin1-3, PIP3, and Akt phosphorylation in addition to the signaling pathways of mTOR complexes inhibited by torin 1 (mTORC1 and mTORC2) or rapamycin (mTORC1). (B) Single-cell quantity dynamics of cells Cavin1-KO (and their corresponding CRISPR management; SI Appendix, Fig. S7D). Statistics are R > 3 for each experiment and n = 7 for management, n = 15 for hypo, n = 18 for hyper. (C) Membrane pressure dynamics of cells Cavin1-KO (and their corresponding CRISPR management; SI Appendix, Fig. S7E). Statistics are n > 4 for each experiment and R = 3 for management, R = 4 for hypo, R = 2 for hyper. (D) Exercise of mTORC1 and mTORC2 underneath hypertonic and hypotonic shock. Panels signify activation (phosphorylation) of S6K1 (p-p70) and Akt (pAkt) and loading controls. Longer timepoints are in SI Appendix, Fig. S7F and corresponding quantification. (E) Single-cell quantity dynamics of cells handled with rapamycin or Torin1 for hypotonic shocks (120 mOsm, circle), isotonic situations (315 mOsm, sq.), and hypertonic shocks (700 mOsm; P/P0 = 2, triangle). Statistics are R > 3 for each experiment. For rapamycin, n = 4 for management, n = 5 for hypo, n = 4 for hyper; for Torin1, n = 10 for management, n = 9 for hypo, n = 7 for hyper. (F) Membrane pressure dynamics of cells handled with rapamycin or Torin1 for an identical shocks as in E. Statistics are n > 4 for each experiment. For rapamycin, R = 2 for management, R = 6 for hypo, R = 4 for hyper; for Torin1, R = 7 for management, R = 15 for hypo, R = 4 for hyper.
Position of mTORCs Exercise throughout Cell Osmotic Response
The fast restoration of cell quantity and pressure throughout hypotonic shocks exhibits that these parameters are tightly and actively regulated by the cell. We studied the position of mTORC1 and mTORC2 on this regulation by following their kinase exercise by means of phosphorylation of p70 and Akt, respectively (Fig. 4A). mTORC2 was rapidly activated underneath hypotonic shock, as beforehand proven (7) (Fig. 4D and SI Appendix, Fig. S7F). We additional noticed that mTORC2 was quickly inhibited underneath hypertonic shock (Fig. 4D). Quite the opposite, mTORC1 exercise remained fixed underneath hypotonic shock and was inhibited underneath hypertonic shock (Fig. 4D) as seen earlier than in ref. 54. The excessive phosphorylation of Akt discovered at 0.1 s (Fig. 4D) was not reproducibly noticed and will come from the truth that these cells had been present process osmotic shocks in chilly buffer, required to dam dephosphorylation. Not like mTORC1, mTORC2 activation had related dynamics underneath hypotonic and hypertonic shocks than quantity and pressure. This correlation advised a extra essential position of mTORC2 than mTORC1 in regulating quantity and pressure response in addition to their coupling. To check this speculation, we pharmacologically inhibited mTORCs and measured cell quantity and pressure responses underneath osmotic shocks. Each mTORC1 and mTORC2 are organized across the kinase mTOR, whose phosphorylation exercise might be totally inhibited utilizing Torin1, which thus inhibits each complexes (55) whereas rapamycin is a particular, partial inhibitor of mTORC1 (56) (Fig. 4A). First, we quantified the kinase exercise of mTORC1 and mTORC2 following rapamycin and Torin1 therapy. We noticed an entire inactivation of each complexes 30 min after Torin1 addition, whereas rapamycin totally inhibited p70 phosphorylation by mTORC1 after 20 min and with out inhibiting mTORC2 (SI Appendix, Fig. S8A) in settlement with ref. 57. We then studied the impact of Torin1 and rapamycin therapies on the cell response to osmotic shocks. Quantity modifications induced by hypotonic shocks had been solely mildly affected by rapamycin, whereas Torin1-treated cells exhibited a considerably lowered quantity peak after hypotonic shocks (Fig. 4E) when in comparison with nontreated cells. Rapamycin-treated cells additionally confirmed a peak of pressure much like that in nontreated cells adopted by a slower restoration of pressure. In Torin1-treated cells, a restricted enhance of pressure, with no restoration, was noticed (Fig. 4F). Each rapamycin- and Torin1-treated cells didn’t present important modifications of their quantity and pressure responses to hypertonic shock compared to nontreated cells (Figs. 1C and 4 E and F and SI Appendix, Fig. S8B), strongly supporting that the cell response to hypertonic shock is actually passive. These outcomes counsel that mTORC2 controls the preliminary quantity/pressure coupling, whereas mTORC2 and mTORC1 are concerned within the long-term restoration of each quantity and pressure. Whereas mTORC complexes are concerned within the regulation of quantity and pressure responses to osmotic shocks and actin within the long-term restoration, components concerned within the preliminary, fast restoration from hypotonic shocks remained unclear. Since actin was depolymerized throughout this section, we postulated that ion transporters stands out as the major actors of this quick restoration.
Position of Ion Transporters throughout Cell Osmotic Response
Exchangers and ion channels are important regulators of cell quantity. VRACs are concerned within the cell response to osmotic shocks whereas NKCC1 and NHE take part within the osmotic steadiness of cells in isotonic situations. Moreover, NHE1 and NKCC1 are interacting with F-actin (58, 59) and mTORCs (Fig. 5A) (24, 60) whereas VRAC has weak interactions with F-actin (61) and its hyperlink with mTORCs remains to be mentioned (62, 63). We used pharmacological inhibitors of channels and transporters concerned in osmotic stress response: the particular LRRC8A channel inhibitor 4-(2-butyl-6,7-dichloro-2-cyclopentylindan-1-on-5-yl)oxybutyric acid (DCPIB) inhibits VRACs, whereas 5-(N-ethyl-N-isopropyl)-Amiloride (EIPA) inhibits NHE exchanger and bumetanide inhibits NKCC1 exchanger (Fig. 5A). After hypotonic shocks, we noticed a gradual affect of medicine from bumetanide to DCPIB on the short-term cell swelling. DCPIB-treated cells had been instantaneously permeabilized upon sturdy hypotonic shock, as seen by the instantaneous labeling of intracellular membranes with CellMask (120 mOsm, Fig. 5B). Cell quantity didn’t change always in milder hypotonic situations (25 and 50% water, Fig. 5C). Against this, cells handled with bumetanide had a smaller however important peak in cell quantity (Fig. 5C), and EIPA-treated cells confirmed no peak instantly after hypotonic shock (Fig. 5C). In EIPA- and bumetanide-treated cells, cell quantity slowly diverged 3 min after shock (Fig. 5C), in line with the position of NHE and NKCC1 in gradual quantity homeostasis (18). All three medicine modified the stress response of cells to hypotonic shocks (Fig. 5D). For bumetanide and EIPA, the response was clearly decoupled from the amount change however whereas EIPA-treated cells had a pressure response equal to that in nontreated cells, in bumetanide-treated cells, pressure didn’t considerably enhance compared to the isotonic management and didn’t get better. Pressure remained fixed for DCPIB-treated cells, completely matching the amount dynamics, however this can be as a result of fast permeabilization of DCPIB-treated cells (Fig. 5B). In all hypertonic situations, not one of the inhibitors examined considerably affected the cell quantity and pressure responses, once more indicating that the hypertonic response is actually passive (Fig. 5 C and D and SI Appendix, Fig. S8C). EIPA has no impact on adaptation from hypertonic shocks whereas EIPA is understood to be an inhibitor of regulatory quantity will increase (RVIs) as a result of we’re a timescale when no quantity enhance had time to happen. Total, these outcomes present that ion transporters that take part within the osmotic steadiness of the cell additionally take part within the coupling between pressure and quantity modifications throughout osmotic shocks and are primarily concerned within the preliminary, giant and fast restoration from hypotonic shocks.

Ion transporters are answerable for the short-term response of cells to osmotic shocks. (A) Illustrations of DCPIB, EIPA, and bumetanide pharmacological results on, respectively, VRACs, NHE, and NKCC1 channels. (B) Confocal photos of DCPIB-treated cells and response underneath hypotonic shock. (Scale bar: 40 µm.) (C) Single-cell quantity dynamics in cells handled with bumetanide, EIPA, and DCPIB for hypotonic shocks (120 mOsm, circle), isotonic situations (315 mOsm, sq.), and hypertonic shocks (700 mOsm; P/P0 = 2, triangle). Statistics are R > 3 for each experiment. For bumetanide, n = 9 for management, n = 66 for hypo, n = 8 for hyper; for EIPA, n = 12 for management, n = 9 for hypo, n = 9 for hyper; for DCPIB, n = 12 for management, n = 31 for hypo, n = 14 for hyper. (D) Membrane pressure dynamics in cells handled with bumetanide, EIPA, and DCPIB for an identical shocks as in C. Statistics are n > 4 for each experiment. For bumetanide, R = 5 for management, R = 11 for hypo, R = 4 for hyper; for EIPA, R = 3 for management, R = 12 for hypo, R = 3 for hyper; for DCPIB, R = 2 for management, R = 6 for hypo, R = 2 for hyper.
Dialogue
Our examine highlights the quantitative relation between cell quantity modifications and cell plasma membrane pressure modifications. We confirmed that cell quantity modifications are primarily as a consequence of cytoplasmic quantity modifications and ensure that cells modulate their quantity in line with the PBVH relation on a bigger scale than beforehand reported (34). We noticed two phases of cell quantity response to osmotic shocks: the short-term response—a couple of seconds after the shock—which was characterised by cell quantity variations in line with the PBVH relation—and the second section—a couple of tens of seconds to minutes after the shock—which we known as the long-term response and was characterised by an uneven restoration. Certainly, cell quantity recovered quick from hypotonic shocks, however recovered slowly from hypertonic shocks. By measuring cell quantity and plasma membrane (PM) pressure for two h after a gentle hypertonic shock (600 mOsm), we noticed that pressure totally recovered, quicker (30 min) than cell quantity (70 min), which recovered solely partially. Reactivation of endocytosis a couple of tens of minutes after the shock (64) may contribute to the quicker pressure restoration by eradicating extra membrane space. Throughout these two phases, we noticed that membrane pressure modifications adopted cell quantity modifications. Within the short-term response, evolution of pressure with quantity modifications was in line with a mannequin based mostly on membrane unfolding. Suits to the mannequin yielded an estimate of the dimensions of the membrane space buffer, i.e., the world saved in ruffles, about 1 to 1.5 occasions the projected space of the cell (maximal space being 2 to 2.5 occasions the preliminary projected space). It additionally offers a worth of the binding free vitality per unit space, which is of the identical order as earlier estimates (51). It additionally enabled inferring the change of pressure in line with the change of strain utilized exterior the cells. This result’s qualitatively maintained through the long-term response, as pressure dynamically evolves with the identical asymmetry as quantity after hypotonic and hypertonic shocks. These outcomes set up that pressure passively follows quantity modifications throughout the whole length of the response and restoration to osmotic shocks. One of many important questions raised by the tight coupling of membrane pressure and quantity modifications throughout osmotic shocks is, What are the determinants of this coupling? After we depleted caveolae, recognized to take part in buffering membrane pressure, we noticed a a lot greater enhance of membrane pressure underneath hypotonic shocks, in line with ref. 6. Cell quantity modifications, then again, had been extra restricted than in parental cells, supporting the concept that depleting caveolae due to this fact removes membrane space buffer, limiting the enlargement of cells throughout osmotic shocks. These outcomes led to the notion that the membrane space buffer is a essential determinant of the coupling between cell quantity and membrane pressure. In additional help of this notion, an inhibitor of the mTOR pathways decoupled pressure and quantity responses within the long-term response, in line with the truth that mTORC1 is proposed to manage cell quantity homeostasis, whereas mTORC2 is proposed to manage cell floor homeostasis (22). Within the short-term response, solely the inhibition of mTORC2 led to a lowered quantity change whereas the stress mildly elevated and didn’t get better. Sturdy mTORC2 inhibition by Torin1 in isotonic medium (SI Appendix, Fig. S9A) didn’t result in PM pressure change however cell quantity is smaller (SI Appendix, Fig. S9B). PM pressure enhance throughout hypotonic shocks is proscribed, suggesting that cells have a excessive membrane space buffer (Figs. 4F and 6A and SI Appendix, Fig. S9B). Constantly, an identical response is discovered when inhibiting NKCC1, which inhibits mTORC2 (24). mTORC2 is likely to be the first signaling advanced coupling cell quantity to pressure as a consequence of its hyperlink to caveolae. Certainly, research spotlight a signaling pathway between caveolae and mTORC2 in mammals (7) much like that between eisosomes and TORC2 in yeast (25), suggesting a typical mechanism of membrane pressure regulation. In mammals, PLD2 interacts with parts of membrane invaginations (clathrin-coated pits and caveolae) and the mTORC2 advanced (7), much like that in Slm1 shuttles between eisosomes and TORC2, to inhibit its exercise when pressure will increase (27). The suggestions loops as mTORC2 regulates the dynamics of caveolin-1 phosphorylation (65) however the hyperlink between caveolae and Akt phosphorylation is underneath debate (66). Additional proof that the membrane space buffer immediately affected the coupling between pressure and quantity got here from learning downstream effectors of mTORC2 and notably the cytoskeleton. When microtubules had been stabilized, the amount of cells elevated (SI Appendix, Fig. S7B) in addition to the osmotically inactive quantity (SI Appendix, Fig. S9C). As a consequence, throughout hypotonic shocks, additional quantity enhance was smaller, in addition to pressure, which didn’t get better (Fig. 3 B and C). These outcomes are appropriate with the notion that elevated preliminary quantity would cut back the membrane space buffer, limiting additional modifications (Fig. 6A). However even when the general response was modified, the coupling between pressure and quantity change was preserved (Figs. 3 B and C and 6B). Altogether, these outcomes help the speculation of laws of cell quantity and pressure unbiased from the regulation of their coupling. The same quantity response to that in taxol-treated cells was obtained with EIPA, because it utterly abrogated the preliminary quantity enhance underneath hypotonic shock. It’s stunning since EIPA is called an inhibitor of RVIs largely examined within the context of mitosis (67) or adherence (68) however, to our information, cell quantity dynamics of EIPA-treated cells present process hypotonic shocks have by no means been examined earlier than. As seen for taxol-treated cells, EIPA-treated cells have bigger cell quantity in isotonic medium (68) (SI Appendix, Fig. S9B) and a bigger osmotically inactive quantity (SI Appendix, Fig. S9D), once more supporting the notion that enhance in preliminary quantity limits additional enhance upon hypotonic shocks. Once more, these outcomes help the concept that greater preliminary quantity reduces the membrane space buffer (SI Appendix, Fig. S9B), limiting additional modifications of quantity and pressure throughout osmotic shocks. Constantly, cells with a smaller preliminary quantity, handled with latrunculin A, jasplakinolide, or nocodazole (SI Appendix, Fig. S7B), bear swelling underneath hypotonic shock with out enhance of their PM pressure (Fig. 3 B and C). That is most likely as a result of their membrane space buffer is elevated (Fig. 6C). Total, our outcomes help the notion that a big extra of membrane is saved in ruffles maintained by the cytoskeleton and that this space buffer units the coupling of cell quantity and membrane pressure modifications throughout osmotic shocks (SI Appendix, Fig. S10). The restoration section is required to revive this huge extra. Any preliminary modification of this membrane extra will affect the coupling of cell membrane pressure and quantity. When the cell quantity dramatically will increase due to hypotonicity, the cell initially responds by depolymerizing the cytoskeleton to drive membrane unfolding, which ends up in a launch of membrane floor space. The preliminary quantity restoration is mediated by means of ion transporters, because the cytoskeleton remains to be disrupted, and finalized with actin repolymerization to refold the membrane, underneath the management of mTOR signaling. Our outcomes present that the coupling between pressure and quantity is actively regulated by the cytoskeleton, ion transporters, and mTOR signaling to keep up a quantitative relation between quantity and pressure effectively described by passive bodily mechanisms. This coupling is regulated by the surplus membrane buffer and is thus partially unbiased of particular regulatory mechanisms of pressure and quantity.

Recapitulative scheme. (A) Within the case of low membrane space buffer, cells have bigger preliminary volumes that stay largely unchanged throughout hypotonic shocks. Membrane ruffles are initially unfolded, which results in greater stretching of the membrane and a rise of PM pressure in resting situations. (B) Within the case of intermediate membrane space buffer, cell volumes enhance underneath hypotonic shocks inflicting the unfolding of membrane ruffles. Membrane stretches and PM pressure will increase in a cell volume-dependent method. (C) Within the case of excessive membrane space buffer, cells have smaller preliminary quantity, thus having additionally extra ruffles and due to this fact cell quantity enhance underneath hypotonic shocks. Cell quantity enhance throughout hypotonic shock is answerable for unfolding the ruffles with out stretching the membrane to its most, due to this fact having restricted enhance of PM pressure.













{kind=link}